

Structural and catalytic effects of proline substitution and surface loop deletion in the extended active site of human carbonic anhydrase II
- PMID: 25683338
- PMCID: PMC4400229
- DOI: 10.1111/febs.13232
Structural and catalytic effects of proline substitution and surface loop deletion in the extended active site of human carbonic anhydrase II
Abstract
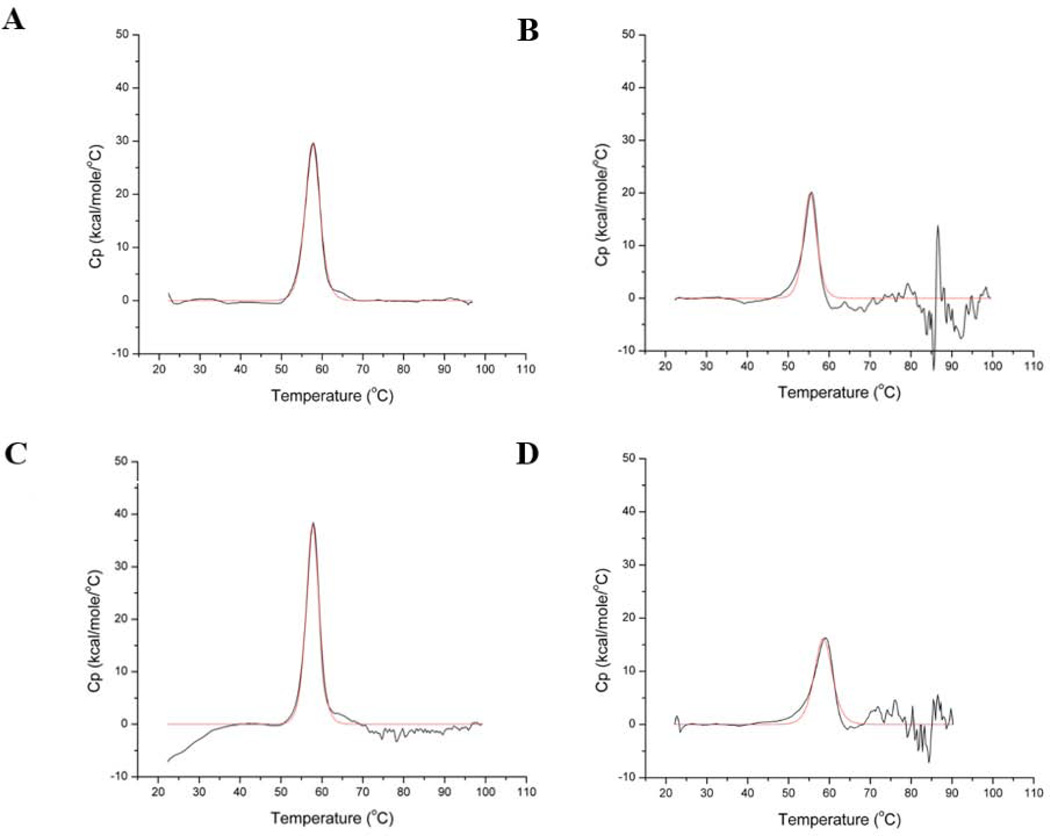
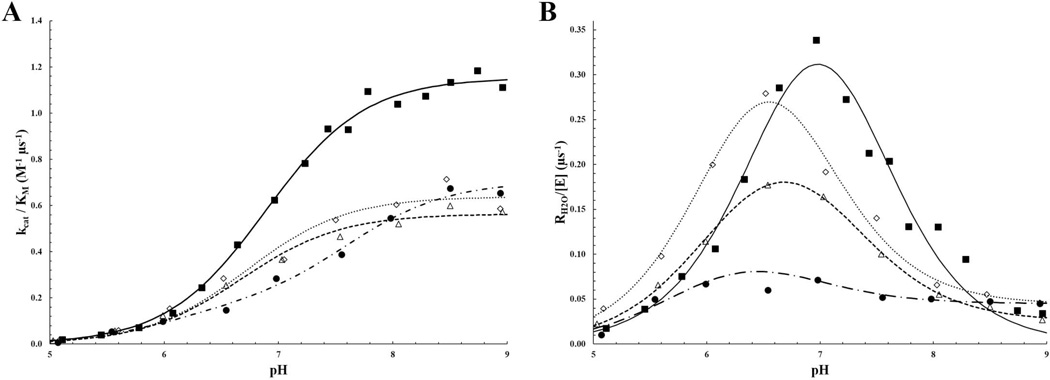
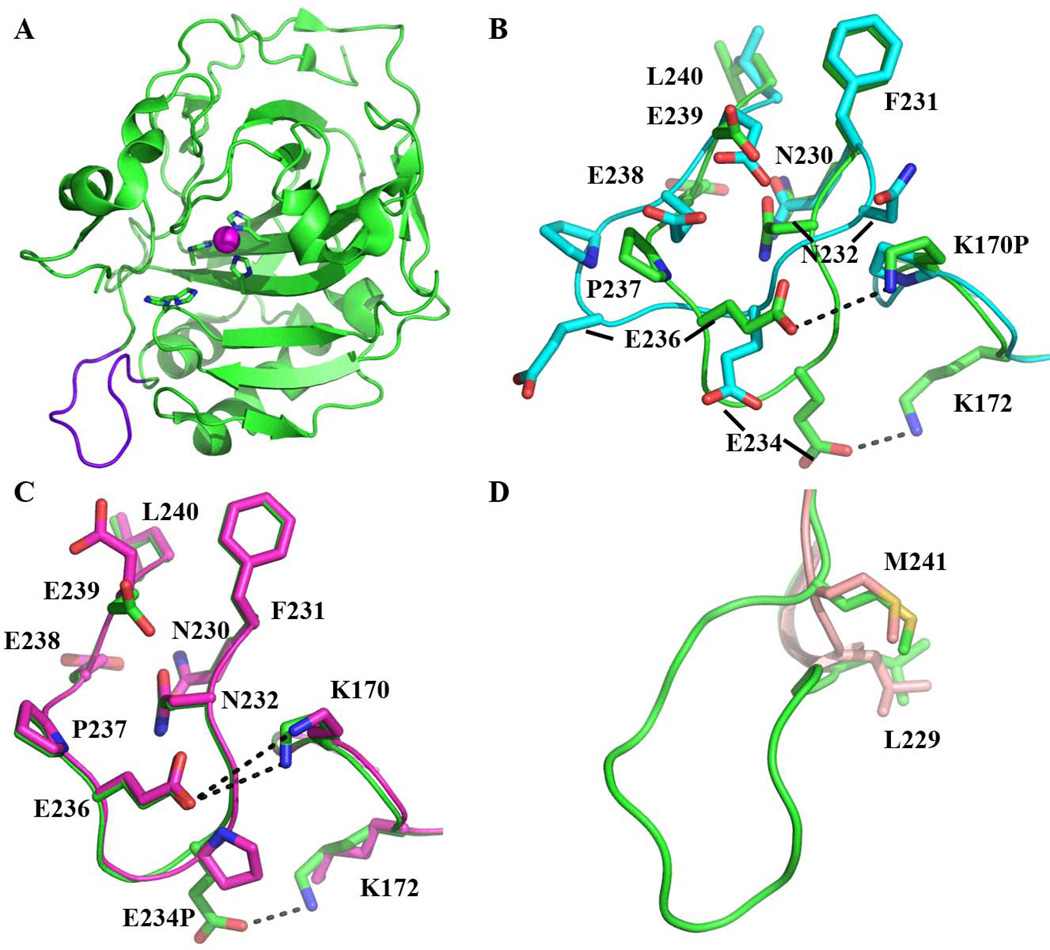
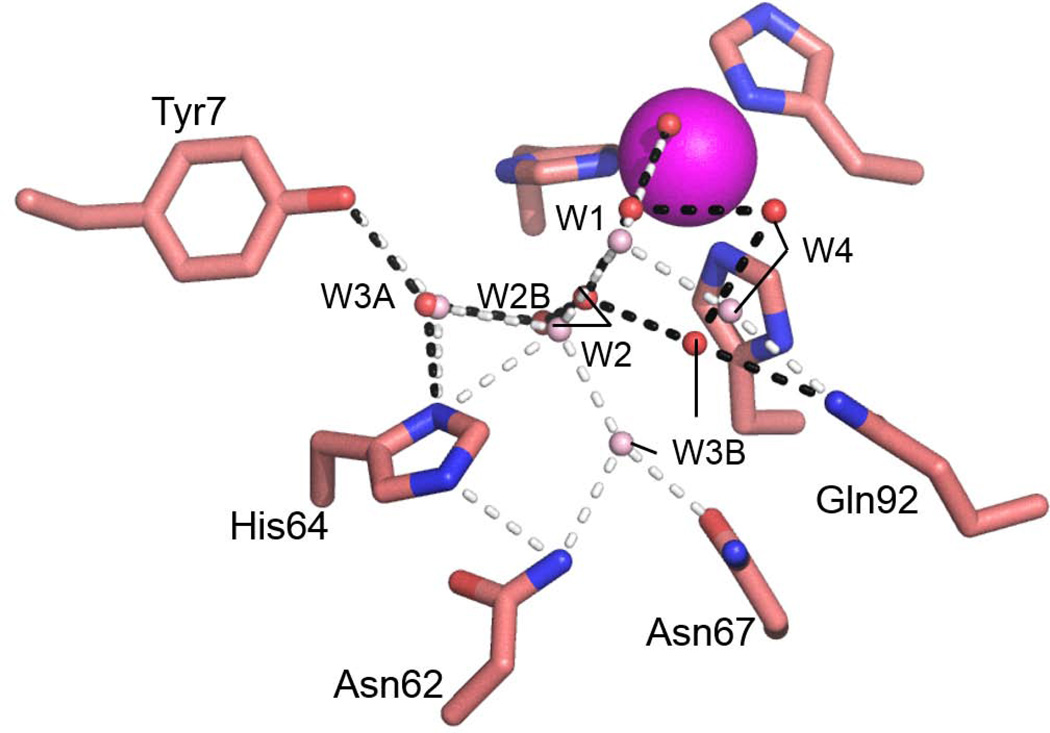
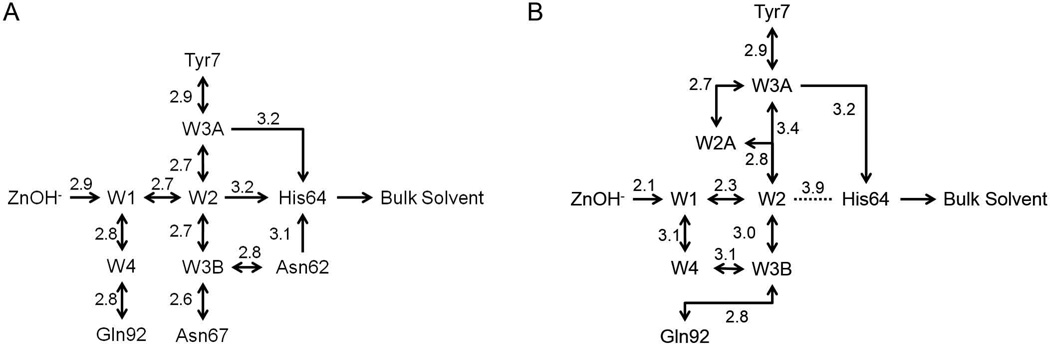
Bioengineering of a thermophilic enzyme starting from a mesophilic scaffold has proven to be a significant challenge, as several stabilizing elements have been proposed to be the foundation of thermal stability, including disulfide bridges, surface loop reduction, ionic pair networks, proline substitutions and aromatic clusters. This study emphasizes the effect of increasing the rigidity of human carbonic anhydrase II (HCA II; EC 4.2.1.1) via incorporation of proline residues at positions 170 and 234, which are located in surface loops that are able to accommodate restrictive main-chain conformations without rearrangement of the surrounding peptide backbone. Additionally, the effect of the compactness of HCA II was examined by deletion of a surface loop (residues 230-240) that had been previously identified as a possible source of thermal stability for the hyperthermophilic carbonic anhydrase isolated from the bacterium Sulfurihydrogenibium yellowstonense YO3AOP1. Differential scanning calorimetry analysis of these HCA II variants revealed that these structural modifications had a minimum effect on the thermal stability of the enzyme, while kinetic studies showed unexpected effects on the catalytic efficiency and proton transfer rates. X-ray crystallographic analysis of these HCA II variants showed that the electrostatic potential and configuration of the highly acidic loop (residues 230-240) play an important role in its high catalytic activity. Based on these observations and previous studies, a picture is emerging of the various components within the general structural architecture of HCA II that are key to stability. These elements may provide blueprints for rational thermal stability engineering of other enzymes.
Database: Structural data have been submitted to the Protein Data Bank under accession numbers 4QK1 (K170P), 4QK2 (E234P) and 4QK3 (Δ230-240).
Keywords: bioengineering; carbonic anhydrase; differential scanning calorimetry; protein crystallography; protein thermal stability.
© 2015 FEBS.
Figures
Similar articles
-
Structural, catalytic and stabilizing consequences of aromatic cluster variants in human carbonic anhydrase II.Arch Biochem Biophys. 2013 Nov 1;539(1):31-7. doi: 10.1016/j.abb.2013.09.001. Epub 2013 Sep 10. Arch Biochem Biophys. 2013. PMID: 24036123 Free PMC article.
-
Kinetic and crystallographic studies of the role of tyrosine 7 in the active site of human carbonic anhydrase II.Arch Biochem Biophys. 2011 Feb 15;506(2):181-7. doi: 10.1016/j.abb.2010.12.004. Epub 2010 Dec 9. Arch Biochem Biophys. 2011. PMID: 21145876 Free PMC article.
-
Speeding up proton transfer in a fast enzyme: kinetic and crystallographic studies on the effect of hydrophobic amino acid substitutions in the active site of human carbonic anhydrase II.Biochemistry. 2007 Mar 27;46(12):3803-13. doi: 10.1021/bi602620k. Epub 2007 Mar 2. Biochemistry. 2007. PMID: 17330962
-
Water networks in fast proton transfer during catalysis by human carbonic anhydrase II.Biochemistry. 2013 Jan 8;52(1):125-31. doi: 10.1021/bi301099k. Epub 2012 Dec 18. Biochemistry. 2013. PMID: 23215152 Free PMC article.
-
Kinetic and structural characterization of thermostabilized mutants of human carbonic anhydrase II.Protein Eng Des Sel. 2012 Jul;25(7):347-55. doi: 10.1093/protein/gzs027. Epub 2012 Jun 12. Protein Eng Des Sel. 2012. PMID: 22691706 Free PMC article.
Cited by
-
Enhancing subtilisin thermostability through a modified normalized B-factor analysis and loop-grafting strategy.J Biol Chem. 2019 Nov 29;294(48):18398-18407. doi: 10.1074/jbc.RA119.010658. Epub 2019 Oct 15. J Biol Chem. 2019. PMID: 31615894 Free PMC article.
-
Catalytic and structural effects of flexible loop deletion in organophosphorus hydrolase enzyme: A thermostability improvement mechanism.J Biosci. 2020;45:54. J Biosci. 2020. PMID: 32345780
-
Engineering de novo disulfide bond in bacterial α-type carbonic anhydrase for thermostable carbon sequestration.Sci Rep. 2016 Jul 7;6:29322. doi: 10.1038/srep29322. Sci Rep. 2016. PMID: 27385052 Free PMC article.
-
Structural and biophysical characterization of the α-carbonic anhydrase from the gammaproteobacterium Thiomicrospira crunogena XCL-2: insights into engineering thermostable enzymes for CO2 sequestration.Acta Crystallogr D Biol Crystallogr. 2015 Aug;71(Pt 8):1745-56. doi: 10.1107/S1399004715012183. Epub 2015 Jul 31. Acta Crystallogr D Biol Crystallogr. 2015. PMID: 26249355 Free PMC article.
-
Activity and anion inhibition studies of the α-carbonic anhydrase from Thiomicrospira crunogena XCL-2 Gammaproteobacterium.Bioorg Med Chem Lett. 2015 Nov 1;25(21):4937-4940. doi: 10.1016/j.bmcl.2015.05.001. Epub 2015 May 6. Bioorg Med Chem Lett. 2015. PMID: 25998503 Free PMC article.
References
-
- Aggarwal M, Boone CD, Kondeti B, McKenna R. Structural annotation of human carbonic anhydrases. J Enzyme Inhib Med Ch. 2013;28:267–277. - PubMed
-
- Boone CD, Pinard M, McKenna R, Silverman D. Catalytic Mechanism of α-Class Carbonic Anhydrases: CO2 Hydration and Proton Transfer. In. In: Frost SC, McKenna R, editors. Carbonic Anhydrase: Mechanism, Regulation, Links to Disease, and Industrial Applications. New York: Springer; 2014. pp. 31–52. - PubMed
-
- Khalifah RG. The carbon dioxide hydration activity of carbonic anhydrase I. Stop-flow kinetic studies on the native human isoenzyme B and C. J Biol Chem. 1971;246:2561–2573. - PubMed
-
- Silverman DN, Lindskog S. The catalytic mechanism of carbonic anhydrase: implications of a rate-limiting protolysis of water. Acc Chem Res. 1988;21:30–36.
-
- Tu CK, Silverman DN, Forsman C, Jonsson BH, Lindskog S. Role of histidine 64 in the catalytic mechanism of human carbonic anhydrase II studied with a site-specific mutant. Biochemistry. 1989;28:7913–7918. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
