

The polymeric mucin Muc5ac is required for allergic airway hyperreactivity
- PMID: 25687754
- PMCID: PMC4333679
- DOI: 10.1038/ncomms7281
The polymeric mucin Muc5ac is required for allergic airway hyperreactivity
Abstract
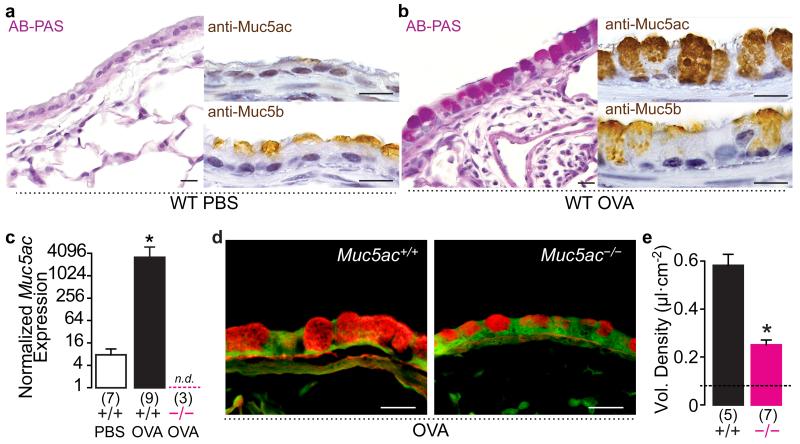
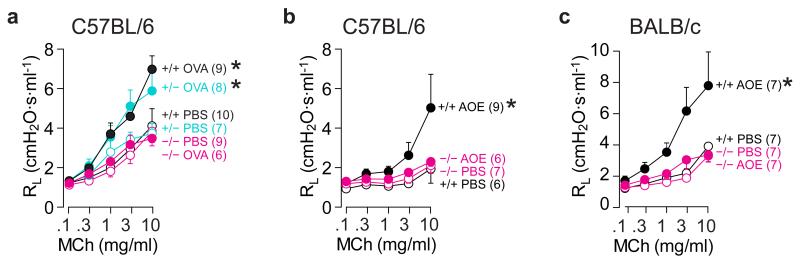
In asthma, airflow obstruction is thought to result primarily from inflammation-triggered airway smooth muscle (ASM) contraction. However, anti-inflammatory and smooth muscle-relaxing treatments are often temporary or ineffective. Overproduction of the mucin MUC5AC is an additional disease feature that, while strongly associated pathologically, is poorly understood functionally. Here we show that Muc5ac is a central effector of allergic inflammation that is required for airway hyperreactivity (AHR) to methacholine (MCh). In mice bred on two well-characterized strain backgrounds (C57BL/6 and BALB/c) and exposed to two separate allergic stimuli (ovalbumin and Aspergillus extract), genetic removal of Muc5ac abolishes AHR. Residual MCh responses are identical to unchallenged controls, and although inflammation remains intact, heterogeneous mucous occlusion decreases by 74%. Thus, whereas inflammatory effects on ASM alone are insufficient for AHR, Muc5ac-mediated plugging is an essential mechanism. Inhibiting MUC5AC may be effective for treating asthma and other lung diseases where it is also overproduced.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
- R01 HL109517/HL/NHLBI NIH HHS/United States
- R01HL070952/HL/NHLBI NIH HHS/United States
- R01 HL097000/HL/NHLBI NIH HHS/United States
- R01HL109517/HL/NHLBI NIH HHS/United States
- R01HL098294/HL/NHLBI NIH HHS/United States
- P30 CA016672/CA/NCI NIH HHS/United States
- R01HL092188/HL/NHLBI NIH HHS/United States
- R21 HL120770/HL/NHLBI NIH HHS/United States
- R01HL119837/HL/NHLBI NIH HHS/United States
- R21ES023384/ES/NIEHS NIH HHS/United States
- P30CA046934/CA/NCI NIH HHS/United States
- P01 HL114457/HL/NHLBI NIH HHS/United States
- UH2HL123442/HL/NHLBI NIH HHS/United States
- UH2 HL123442/HL/NHLBI NIH HHS/United States
- P30CA016672/CA/NCI NIH HHS/United States
- R01HL097000/HL/NHLBI NIH HHS/United States
- R21 ES023384/ES/NIEHS NIH HHS/United States
- R01DK097075/DK/NIDDK NIH HHS/United States
- R01 HL098294/HL/NHLBI NIH HHS/United States
- R21HL120770/HL/NHLBI NIH HHS/United States
- P30 CA046934/CA/NCI NIH HHS/United States
- R01 HL080396/HL/NHLBI NIH HHS/United States
- R01 HL070952/HL/NHLBI NIH HHS/United States
- P01HL114457/HL/NHLBI NIH HHS/United States
- R01HL080396/HL/NHLBI NIH HHS/United States
- R01 DK097075/DK/NIDDK NIH HHS/United States
- R01 HL097163/HL/NHLBI NIH HHS/United States
- R33 HL120770/HL/NHLBI NIH HHS/United States
- R01 HL129795/HL/NHLBI NIH HHS/United States
- R01 HL092188/HL/NHLBI NIH HHS/United States
- R01 HL119837/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
