

Genetic basis for metabolism of methylated sulfur compounds in Methanosarcina species
- PMID: 25691524
- PMCID: PMC4372740
- DOI: 10.1128/JB.02605-14
Genetic basis for metabolism of methylated sulfur compounds in Methanosarcina species
Abstract
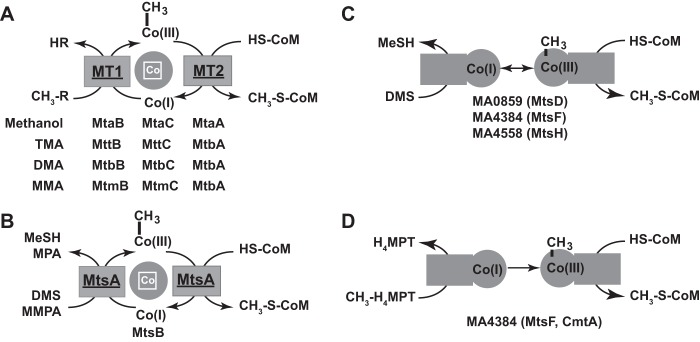
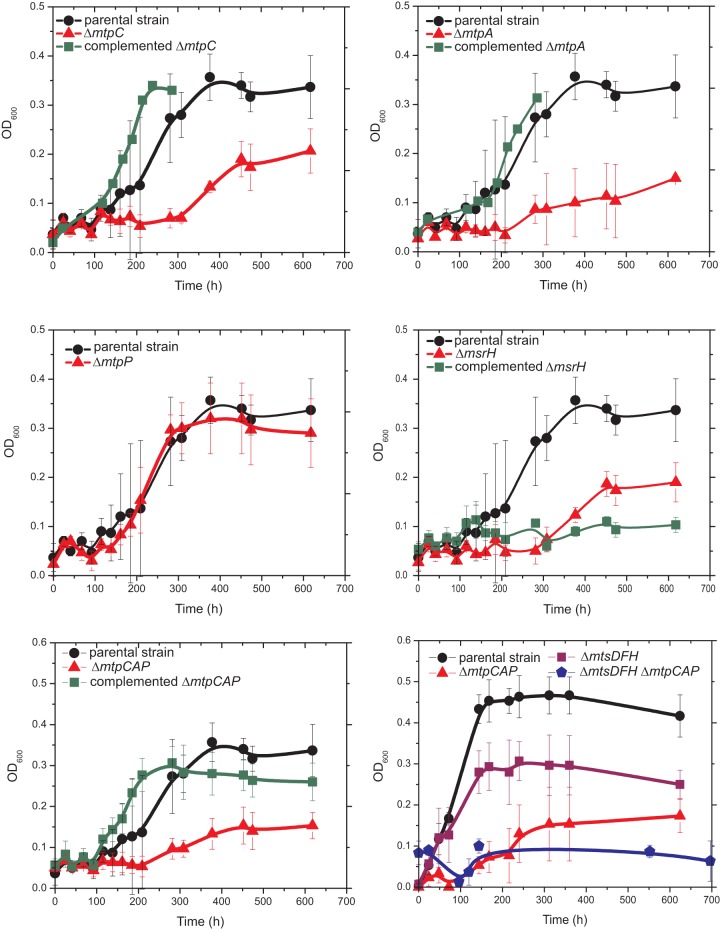
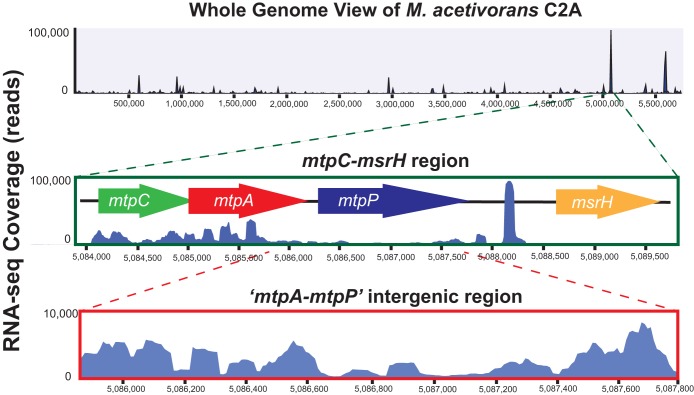
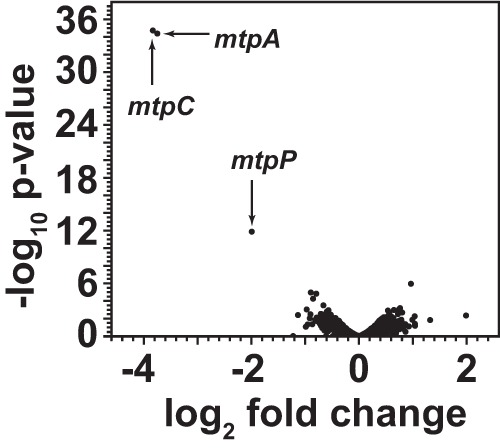
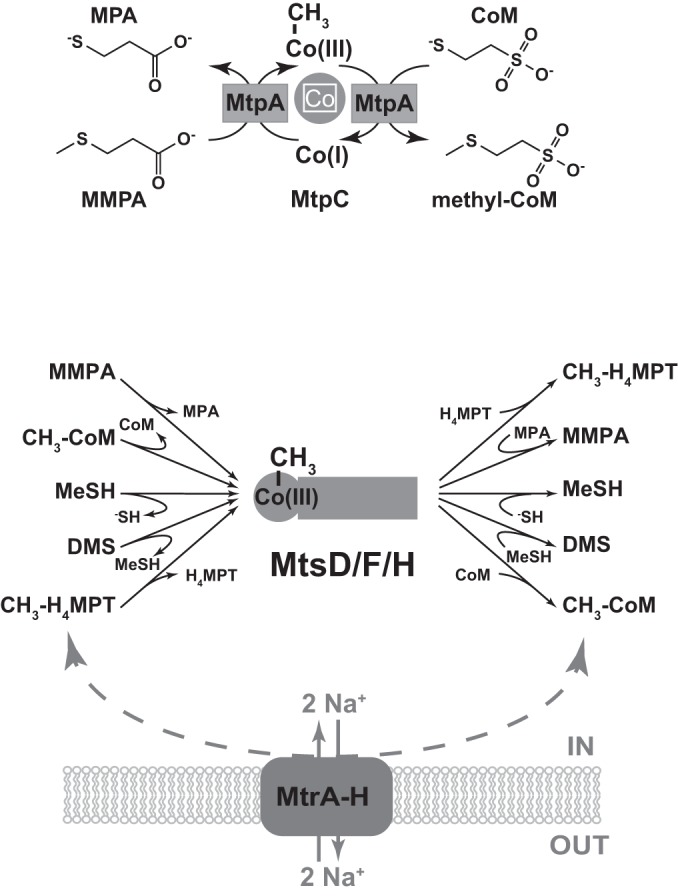
Methanosarcina acetivorans uses a variety of methylated sulfur compounds as carbon and energy sources. Previous studies implicated the mtsD, mtsF, and mtsH genes in catabolism of dimethylsulfide, but the genes required for use of other methylsulfides have yet to be established. Here, we show that a four-gene locus, designated mtpCAP-msrH, is specifically required for growth on methylmercaptopropionate (MMPA). The mtpC, mtpA, and mtpP genes encode a putative corrinoid protein, a coenzyme M (CoM) methyltransferase, and a major facilitator superfamily (MFS) transporter, respectively, while msrH encodes a putative transcriptional regulator. Mutants lacking mtpC or mtpA display a severe growth defect in MMPA medium but are unimpaired during growth on other substrates. The mtpCAP genes comprise a transcriptional unit that is highly and specifically upregulated during growth on MMPA, whereas msrH is monocistronic and constitutively expressed. Mutants lacking msrH fail to transcribe mtpCAP and grow poorly in MMPA medium, consistent with the assignment of its product as a transcriptional activator. The mtpCAP-msrH locus is conserved in numerous marine methanogens, including eight Methanosarcina species that we showed are capable of growth on MMPA. Mutants lacking the mtsD, mtsF, and mtsH genes display a 30% reduction in growth yield when grown on MMPA, suggesting that these genes play an auxiliary role in MMPA catabolism. A quadruple ΔmtpCAP ΔmtsD ΔmtsF ΔmtsH mutant strain was incapable of growth on MMPA. Reanalysis of mtsD, mtsF, and mtsH mutants suggests that the preferred substrate for MtsD is dimethylsulfide, while the preferred substrate for MtsF is methanethiol.
Importance: Methylated sulfur compounds play pivotal roles in the global sulfur and carbon cycles and contribute to global temperature homeostasis. Although the degradation of these molecules by aerobic bacteria has been well studied, relatively little is known regarding their fate in anaerobic ecosystems. In this study, we identify the genetic basis for metabolism of methylmercaptopropionate, dimethylsulfide, and methanethiol by strictly anaerobic methanogens of the genus Methanosarcina. These data will aid the development of predictive sulfur cycle models and enable molecular ecological approaches for the study of methylated sulfur metabolism in anaerobic ecosystems.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
