

Fission yeast Cdk7 controls gene expression through both its CAK and C-terminal domain kinase activities
- PMID: 25691663
- PMCID: PMC4387208
- DOI: 10.1128/MCB.00024-15
Fission yeast Cdk7 controls gene expression through both its CAK and C-terminal domain kinase activities
Abstract
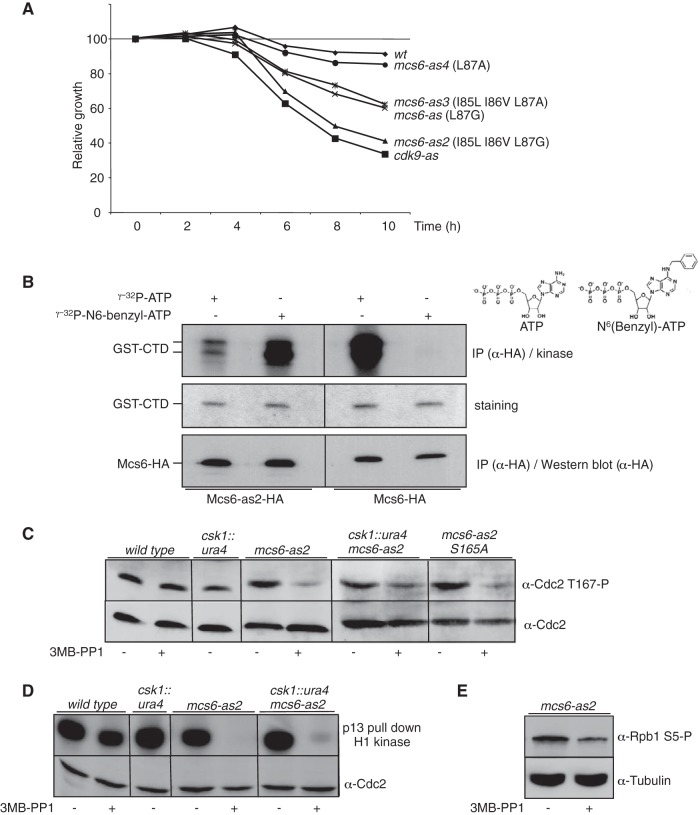
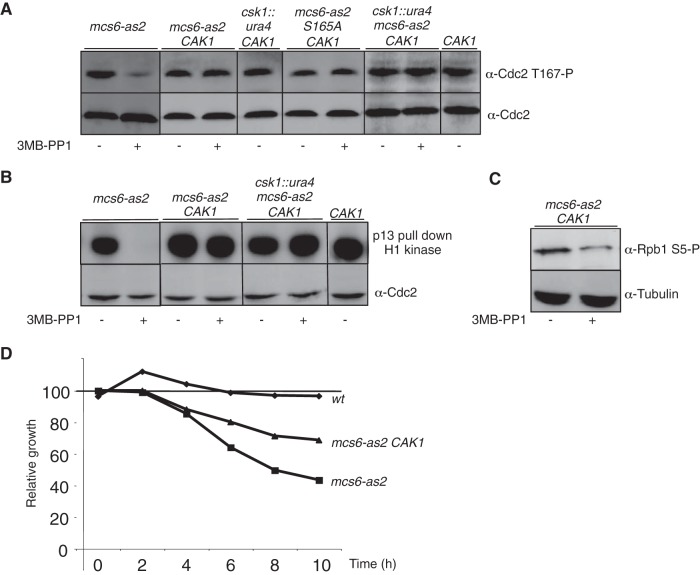
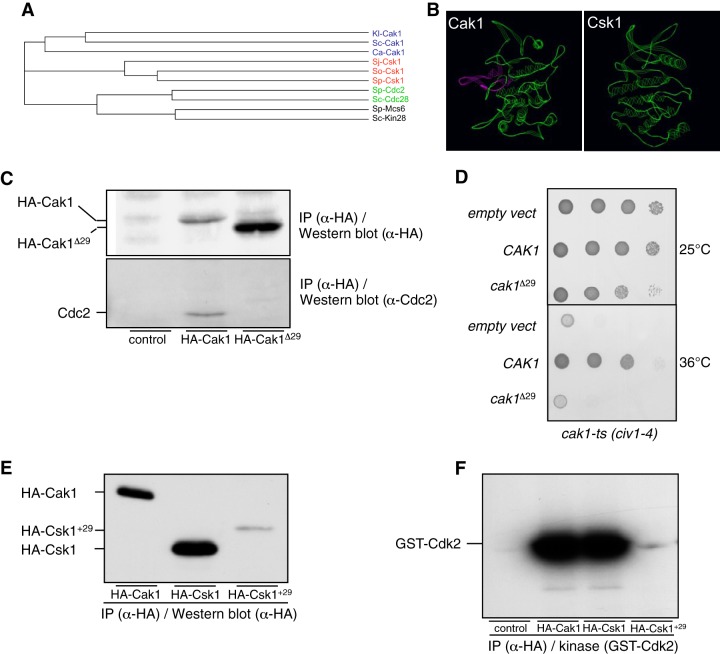
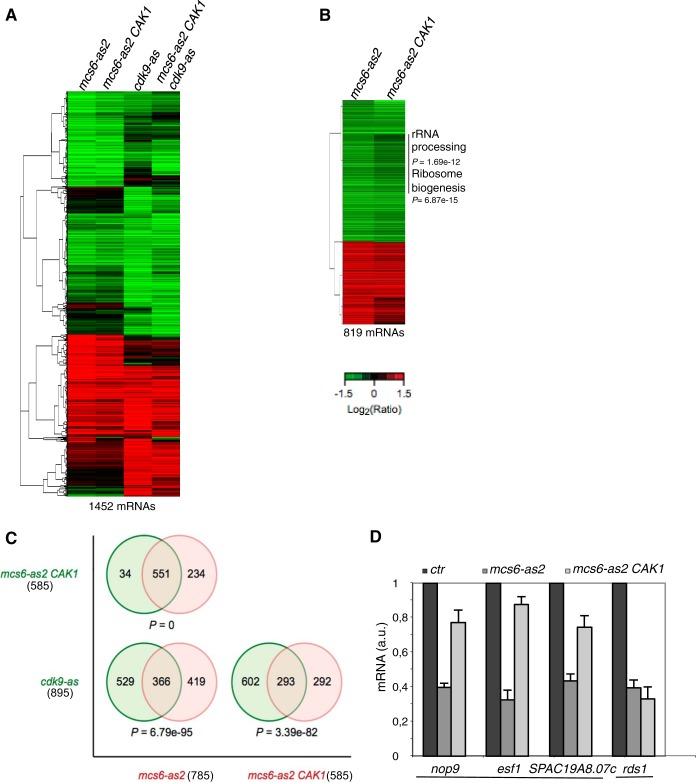
Cyclin-dependent kinase (Cdk) activation and RNA polymerase II transcription are linked by the Cdk7 kinase, which phosphorylates Cdks as a trimeric Cdk-activating kinase (CAK) complex, and serine 5 within the polymerase II (Pol II) C-terminal domain (CTD) as transcription factor TFIIH-bound CAK. However, the physiological importance of integrating these processes is not understood. Besides the Cdk7 ortholog Mcs6, fission yeast possesses a second CAK, Csk1. The two enzymes have been proposed to act redundantly to activate Cdc2. Using an improved analogue-sensitive Mcs6-as kinase, we show that Csk1 is not a relevant CAK for Cdc2. Further analyses revealed that Csk1 lacks a 20-amino-acid sequence required for its budding yeast counterpart, Cak1, to bind Cdc2. Transcriptome profiling of the Mcs6-as mutant in the presence or absence of the budding yeast Cak1 kinase, in order to uncouple the CTD kinase and CAK activities of Mcs6, revealed an unanticipated role of the CAK branch in the transcriptional control of the cluster of genes implicated in ribosome biogenesis and cell growth. The analysis of a Cdc2 CAK site mutant confirmed these data. Our data show that the Cdk7 kinase modulates transcription through its well-described RNA Pol II CTD kinase activity and also through the Cdc2-activating kinase activity.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous