

Ancient dynamin segments capture early stages of host-mitochondrial integration
- PMID: 25691734
- PMCID: PMC4352829
- DOI: 10.1073/pnas.1407163112
Ancient dynamin segments capture early stages of host-mitochondrial integration
Abstract
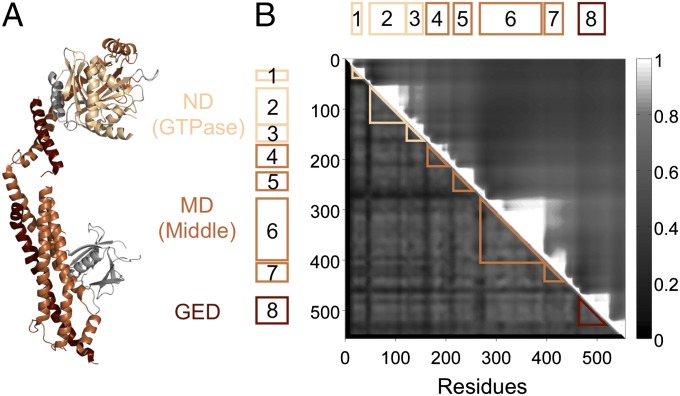
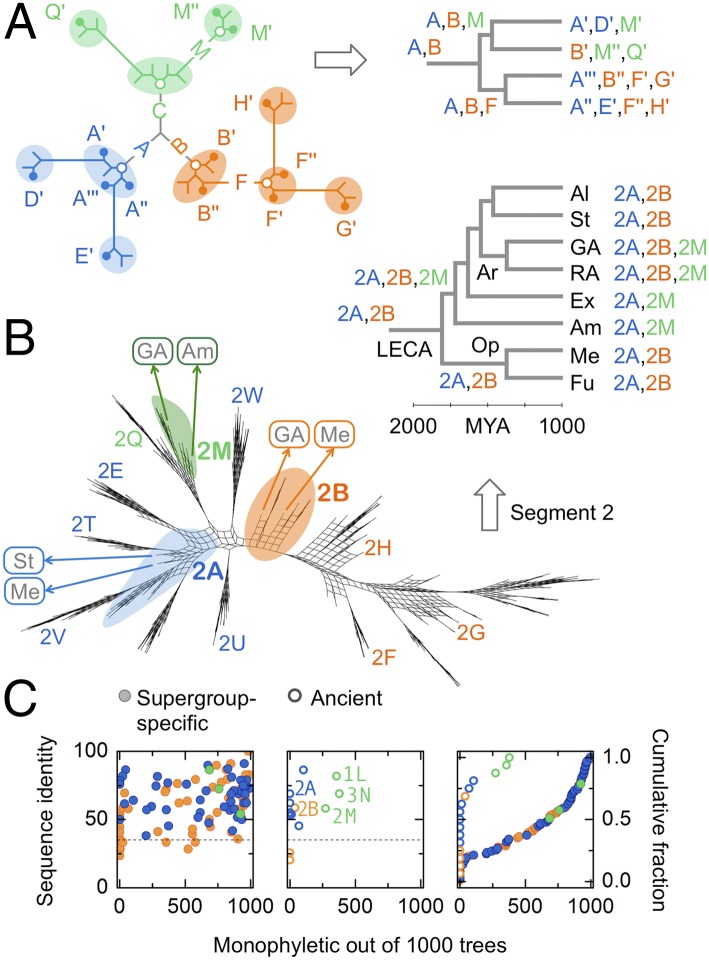
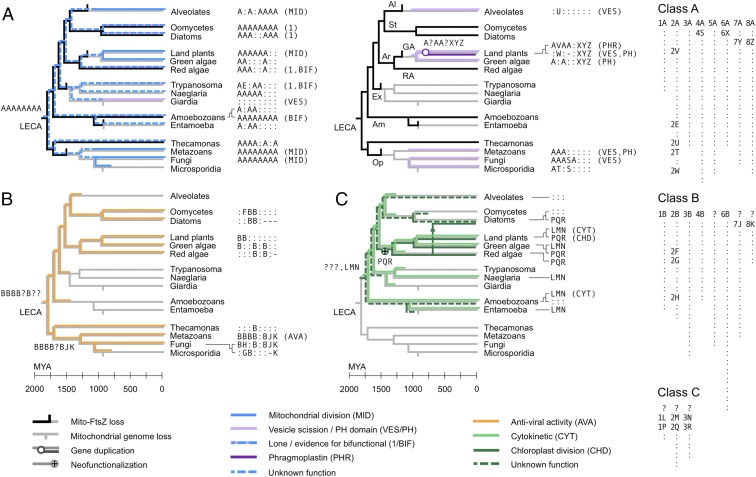
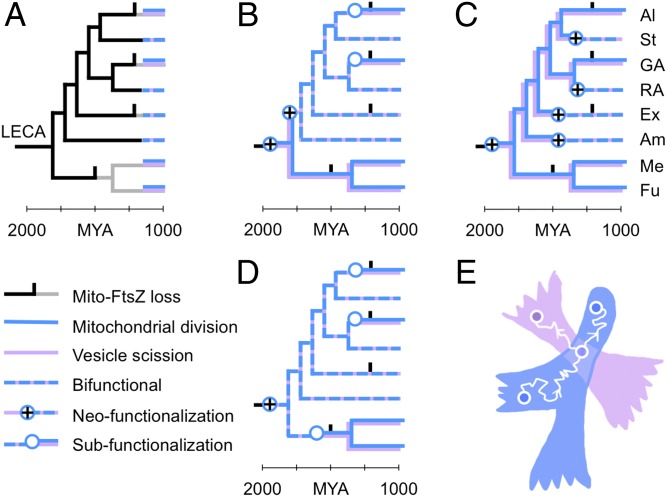
Eukaryotic cells use dynamins-mechano-chemical GTPases--to drive the division of endosymbiotic organelles. Here we probe early steps of mitochondrial and chloroplast endosymbiosis by tracing the evolution of dynamins. We develop a parsimony-based phylogenetic method for protein sequence reconstruction, with deep time resolution. Using this, we demonstrate that dynamins diversify through the punctuated transformation of sequence segments on the scale of secondary-structural elements. We find examples of segments that have remained essentially unchanged from the 1.8-billion-y-old last eukaryotic common ancestor to the present day. Stitching these together, we reconstruct three ancestral dynamins: The first is nearly identical to the ubiquitous mitochondrial division dynamins of extant eukaryotes, the second is partially preserved in the myxovirus-resistance--like dynamins of metazoans, and the third gives rise to the cytokinetic dynamins of amoebozoans and plants and to chloroplast division dynamins. The reconstructed sequences, combined with evolutionary models and published functional data, suggest that the ancestral mitochondrial division dynamin also mediated vesicle scission. This bifunctional protein duplicated into specialized mitochondrial and vesicle variants at least three independent times--in alveolates, green algae, and the ancestor of fungi and metazoans-accompanied by the loss of the ancient prokaryotic mitochondrial division protein FtsZ. Remarkably, many extant species that retain FtsZ also retain the predicted ancestral bifunctional dynamin. The mitochondrial division apparatus of such organisms, including amoebozoans, red algae, and stramenopiles, seems preserved in a near-primordial form.
Keywords: FtsZ; dynamin; eukaryote evolution; mitochondria; vesicles.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Williams TA, Foster PG, Cox CJ, Embley TM. An archaeal origin of eukaryotes supports only two primary domains of life. Nature. 2013;504(7479):231–236. - PubMed
-
- Keeling PJ. Diversity and evolutionary history of plastids and their hosts. Am J Bot. 2004;91(10):1481–1493. - PubMed
-
- Osteryoung KW, Nunnari J. The division of endosymbiotic organelles. Science. 2003;302(5651):1698–1704. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
