

STRUCTURAL AND FUNCTIONAL CONSEQUENCES OF CIRCULAR PERMUTATION ON THE ACTIVE SITE OF OLD YELLOW ENZYME
- PMID: 25692074
- PMCID: PMC4327928
- DOI: 10.1021/cs501702k
STRUCTURAL AND FUNCTIONAL CONSEQUENCES OF CIRCULAR PERMUTATION ON THE ACTIVE SITE OF OLD YELLOW ENZYME
Abstract
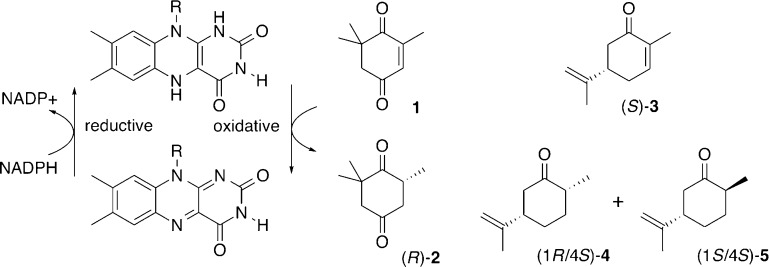
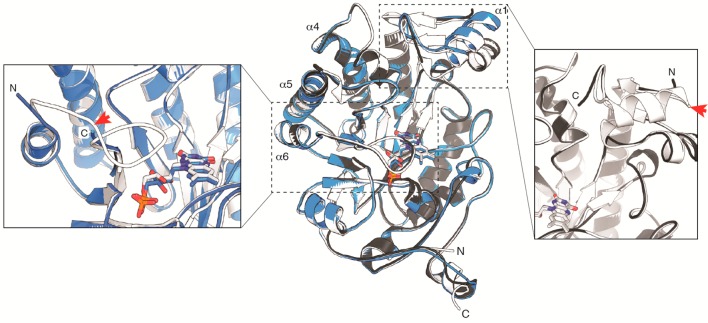
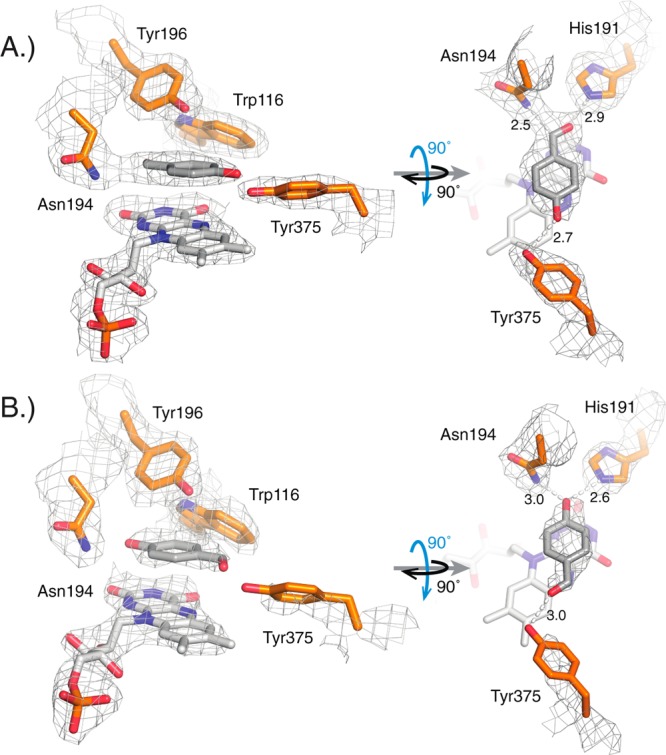
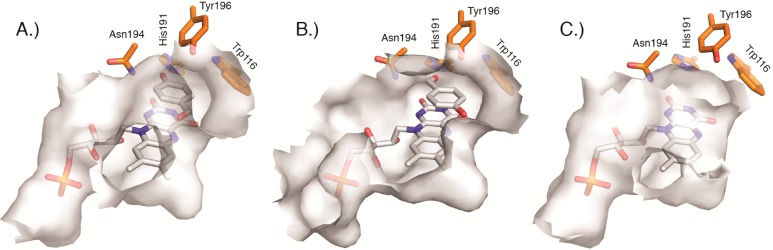
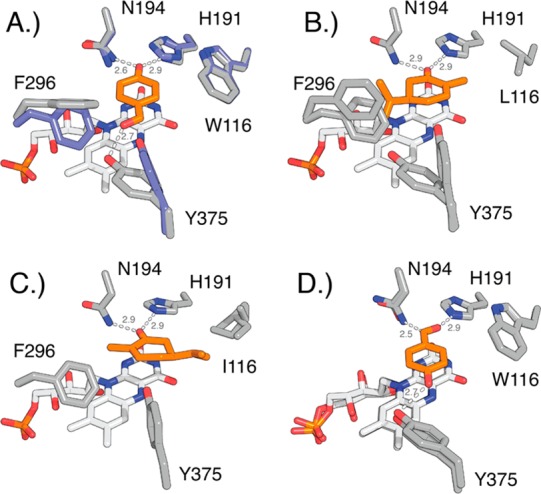
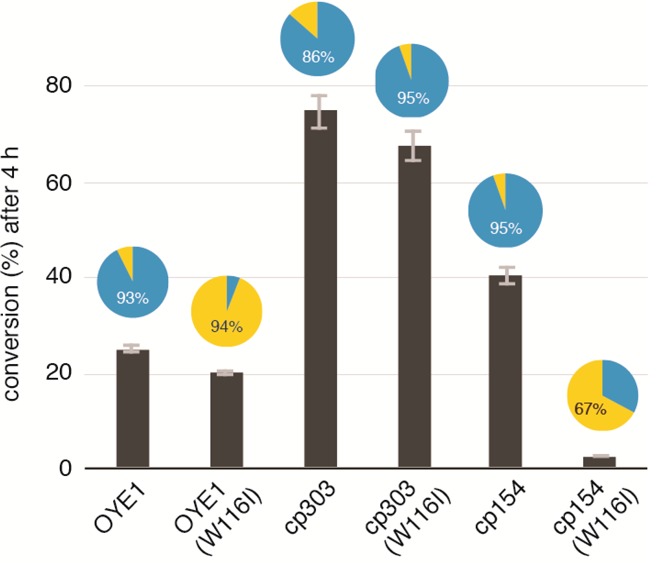
Circular permutation of the NADPH-dependent oxidoreductase Old Yellow Enzyme from Saccharomyces pastorianus (OYE1) can significantly enhance the enzyme's catalytic performance. Termini relocation into four regions of the protein (sectors I-IV) near the active site has proven effective in altering enzyme function. To better understand the structural consequences and rationalize the observed functional gains in these OYE1 variants, we selected representatives from sectors I-III for further characterization by biophysical methods and X-ray crystallography. These investigations not only show trends in enzyme stability and quaternary structure as a function of termini location, but also provide a possible explanation for the catalytic gains in our top-performing OYE variant (new N-terminus at residue 303; sector III). Crystallographic analysis indicates that termini relocation into sector III affects the loop β6 region (amino acid positions: 290-310) of OYE1 which forms a lid over the active site. Peptide backbone cleavage greatly enhances local flexibility, effectively converting the loop into a tether and consequently increasing the environmental exposure of the active site. Interestingly, such active site remodeling does not negatively impact the enzyme's activity and stereoselectivity, nor does it perturb the conformation of other key active site residues with the exception of Y375. These observations were confirmed in truncation experiments, deleting all residues of the loop β6 region in our OYE variant. Intrigued by the finding that circular permutation leaves most of the key catalytic residues unchanged, we also tested OYE permutants for possible additive or synergistic effects of amino acid substitutions. Distinct functional changes in these OYE variants were detected upon mutations at W116, known in native OYE1 to cause inversion of diastereo-selectivity for (S)-carvone reduction. Our findings demonstrate the contribution of loop β6 toward determining the stereoselectivity of OYE1, an important insight for future OYE engineering efforts.
Keywords: Old Yellow Enzyme; X-ray crystallography; biocatalysis; circular permutation; oxidoreductases; protein engineering.
Figures
References
-
- Kawai Y.; Inaba Y.; Tokitoh N. Tetrahedron-Asymmetry 2001, 12, 309–31810.1016/S0957-4166(01)00029-5. - DOI
-
- Swiderska M. A.; Stewart J. D. J. Mol. Catal., B: Enzymol. 2006, 42, 52–5410.1016/j.molcatb.2006.06.023. - DOI
-
- Hall M.; Stueckler C.; Hauer B.; Stuermer R.; Friedrich T.; Breuer M.; Kroutil W.; Faber K. Eur. J. Org. Chem. 2008, 1511–151610.1002/ejoc.200701208. - DOI
-
- Kosjek B.; Fleitz F. J.; Dormer P. G.; Kuethe J. T.; Devine P. N. Tetrahedron-Asymmetry 2008, 19, 1403–140610.1016/j.tetasy.2008.05.023. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources