

A cnidarian homologue of an insect gustatory receptor functions in developmental body patterning
- PMID: 25692633
- PMCID: PMC4374167
- DOI: 10.1038/ncomms7243
A cnidarian homologue of an insect gustatory receptor functions in developmental body patterning
Abstract
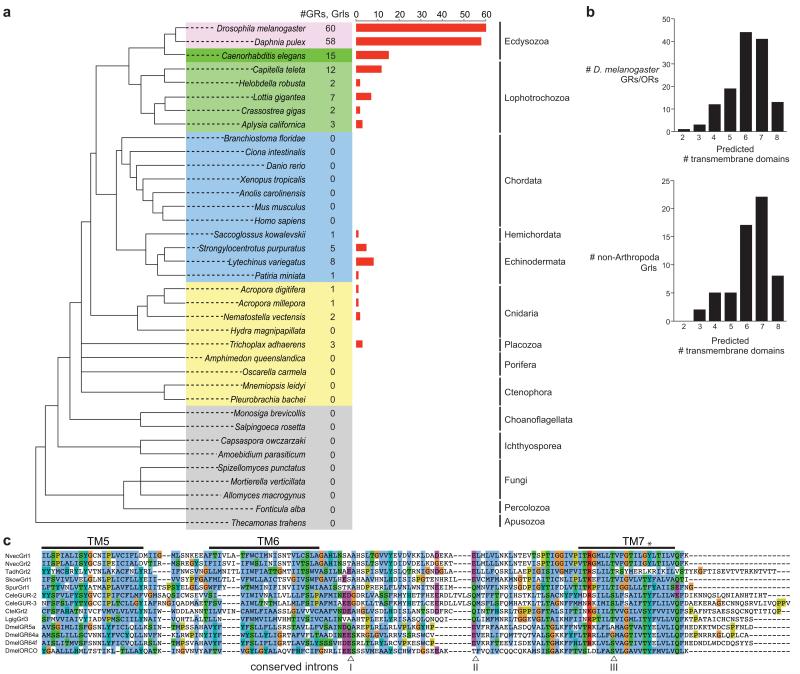
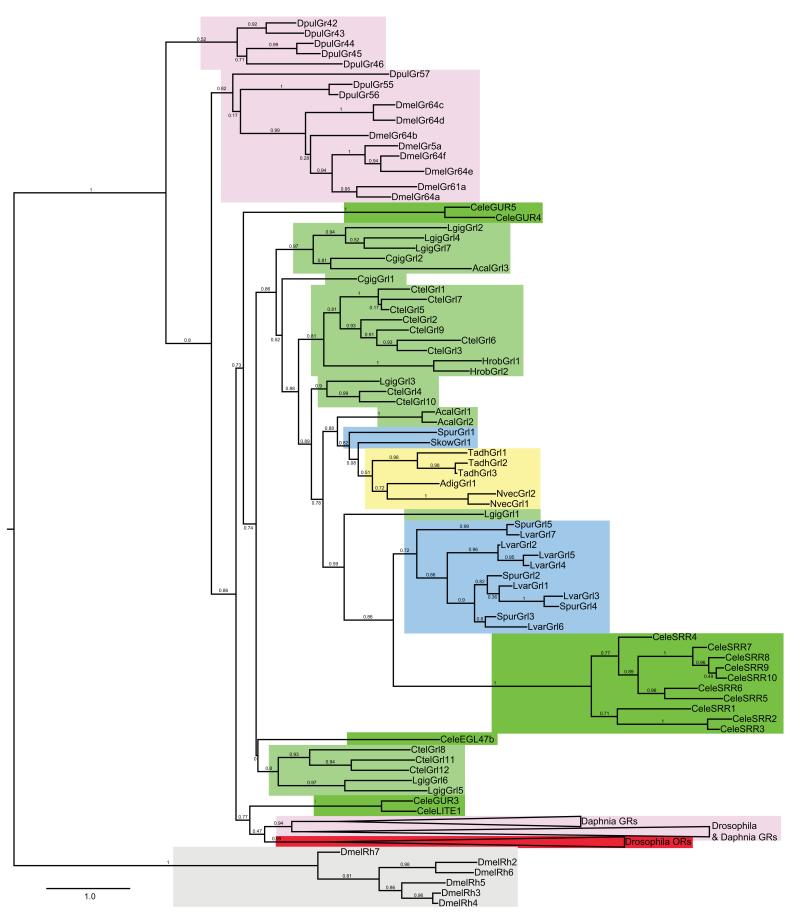
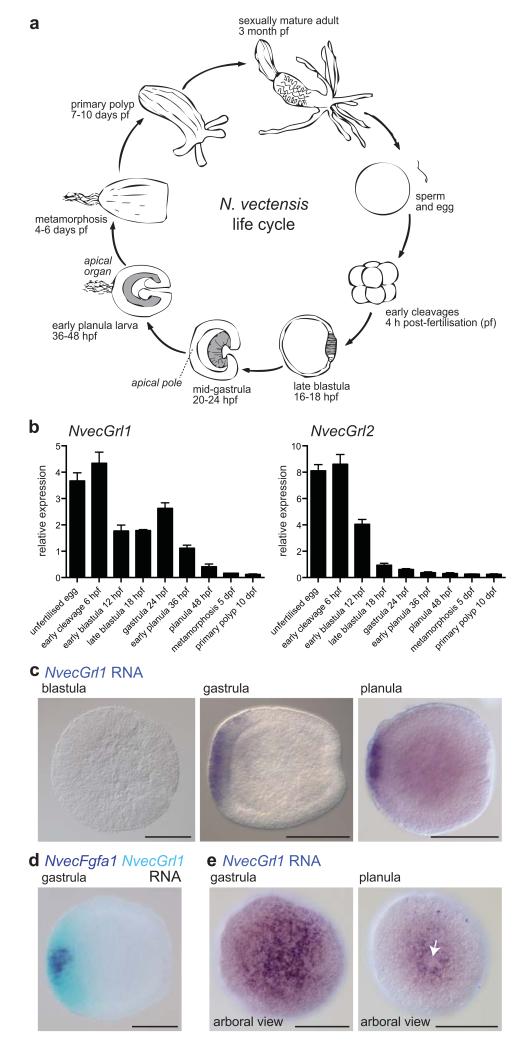
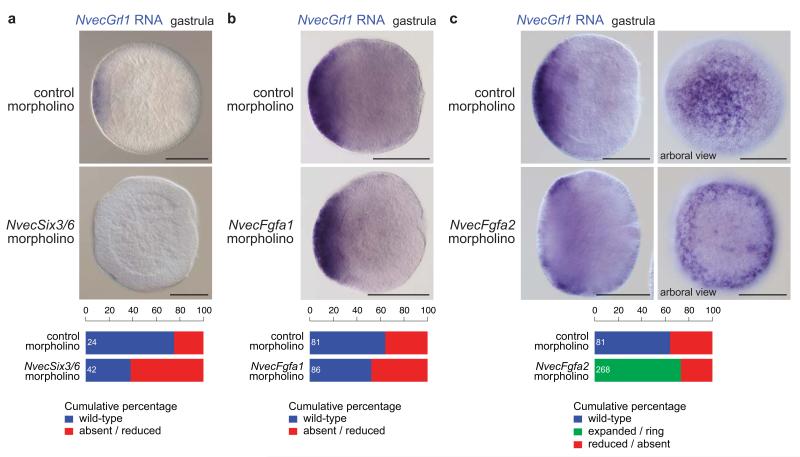
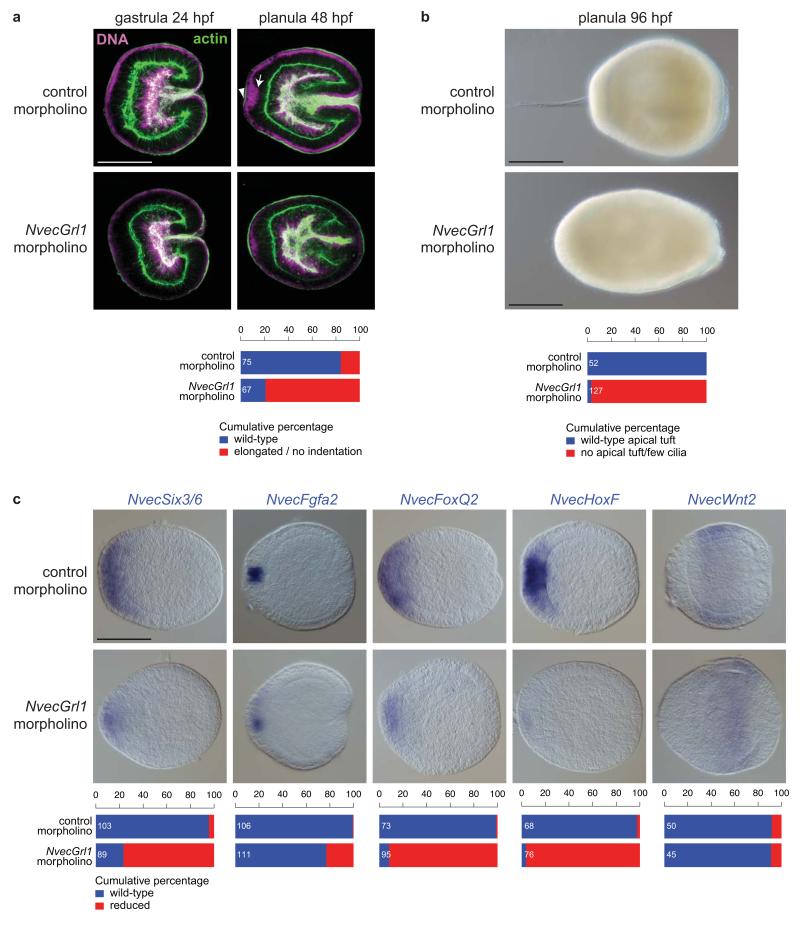
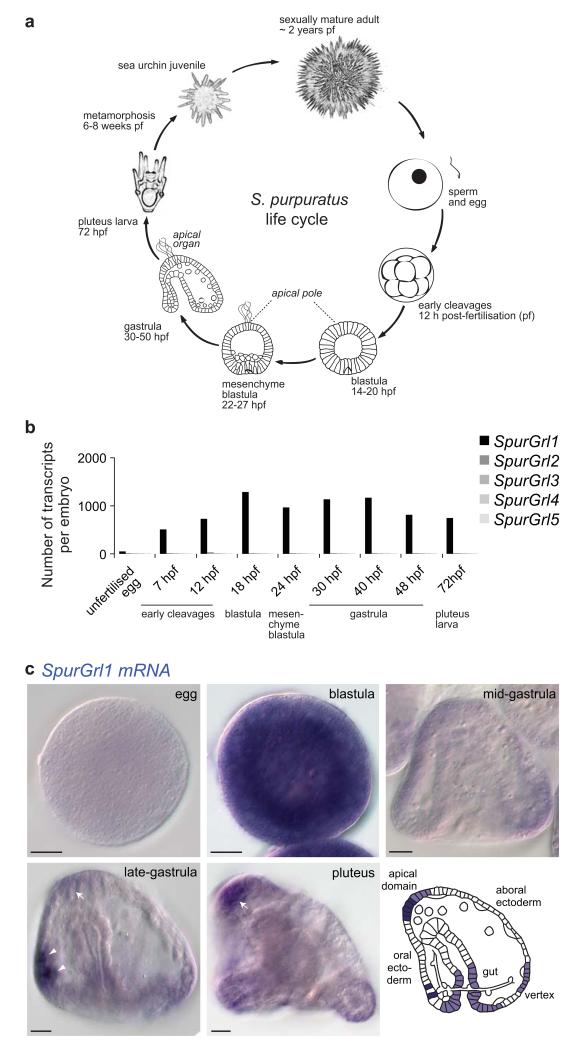
Insect gustatory and odorant receptors (GRs and ORs) form a superfamily of novel transmembrane proteins, which are expressed in chemosensory neurons that detect environmental stimuli. Here we identify homologues of GRs (Gustatory receptor-like (Grl) genes) in genomes across Protostomia, Deuterostomia and non-Bilateria. Surprisingly, two Grls in the cnidarian Nematostella vectensis, NvecGrl1 and NvecGrl2, are expressed early in development, in the blastula and gastrula, but not at later stages when a putative chemosensory organ forms. NvecGrl1 transcripts are detected around the aboral pole, considered the equivalent to the head-forming region of Bilateria. Morpholino-mediated knockdown of NvecGrl1 causes developmental patterning defects of this region, leading to animals lacking the apical sensory organ. A deuterostome Grl from the sea urchin Strongylocentrotus purpuratus displays similar patterns of developmental expression. These results reveal an early evolutionary origin of the insect chemosensory receptor family and raise the possibility that their ancestral role was in embryonic development.
Figures
References
-
- Hallem EA, Dahanukar A, Carlson JR. Insect odor and taste receptors. Annu. Rev. Entomol. 2006;51:113–135. - PubMed
-
- Benton R. Chemical sensing in Drosophila. Curr. Opin. Neurobiol. 2008;18:357–363. - PubMed
-
- Nei M, Niimura Y, Nozawa M. The evolution of animal chemosensory receptor gene repertoires: roles of chance and necessity. Nat. Rev. Genet. 2008;9:951–963. - PubMed
-
- Vosshall LB, Stocker RF. Molecular Architecture of Smell and Taste in Drosophila. Annu. Rev. Neurosci. 2007;30:505–533. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
