

Multiplex CRISPR/Cas9-based genome editing for correction of dystrophin mutations that cause Duchenne muscular dystrophy
- PMID: 25692716
- PMCID: PMC4335351
- DOI: 10.1038/ncomms7244
Multiplex CRISPR/Cas9-based genome editing for correction of dystrophin mutations that cause Duchenne muscular dystrophy
Abstract
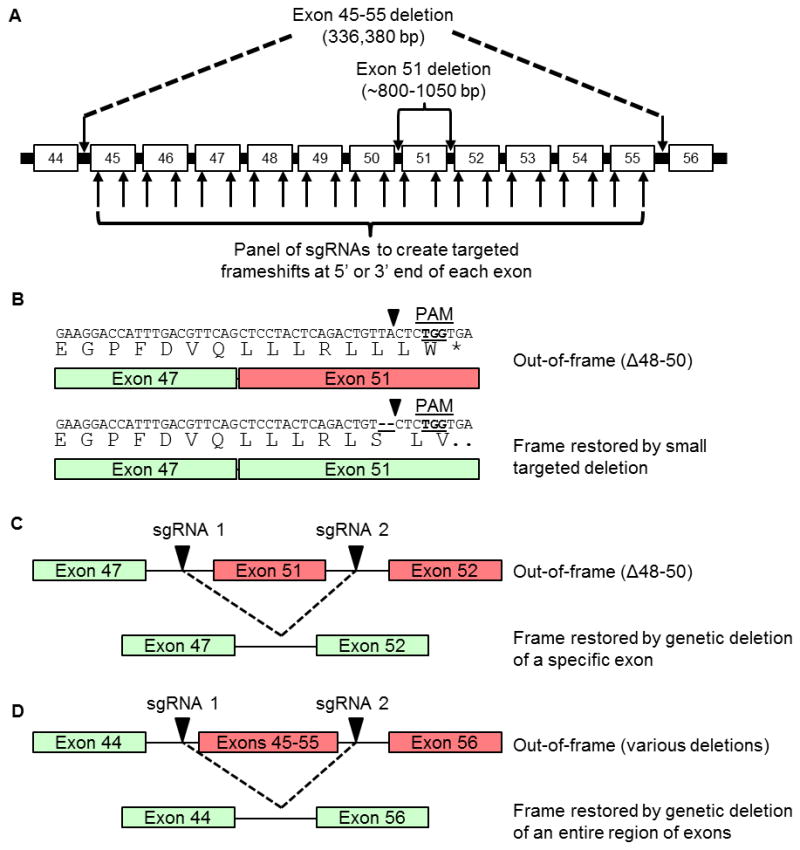
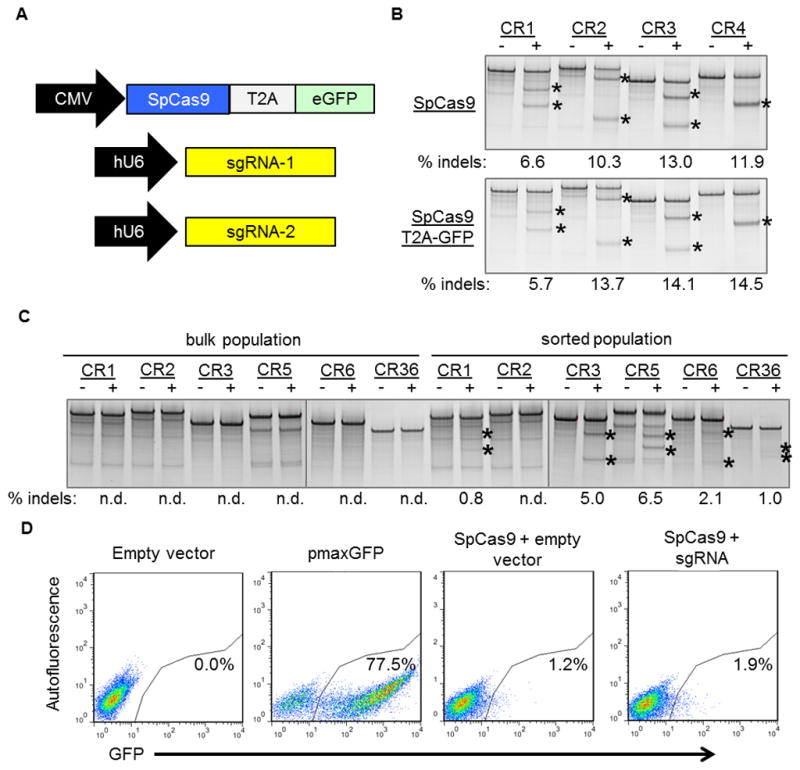
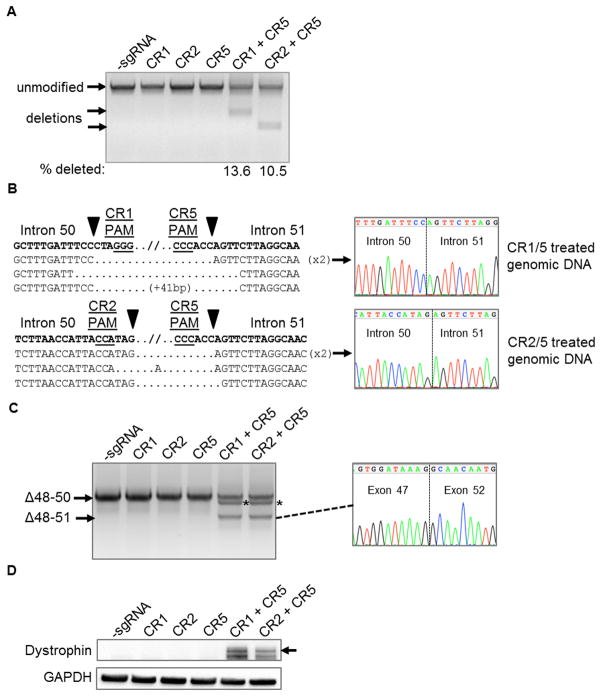
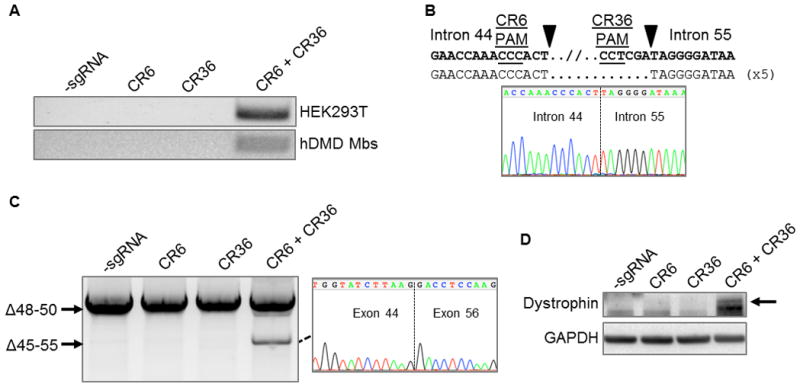
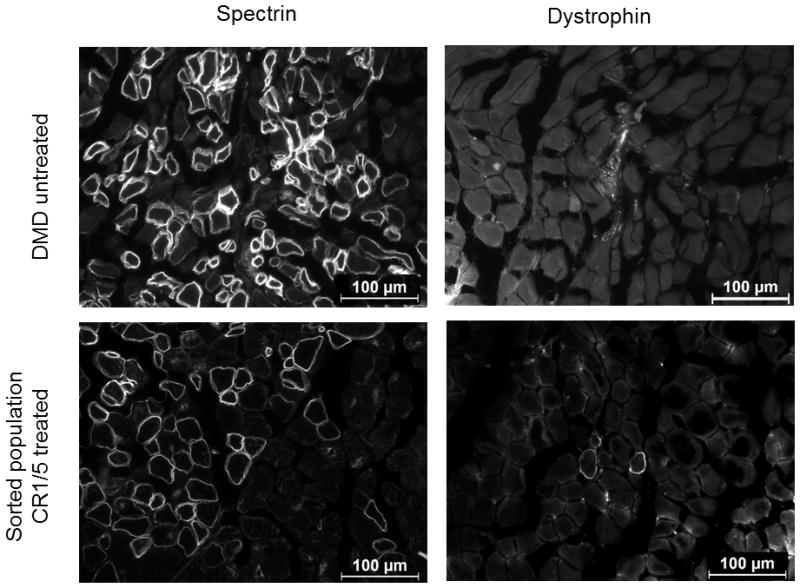
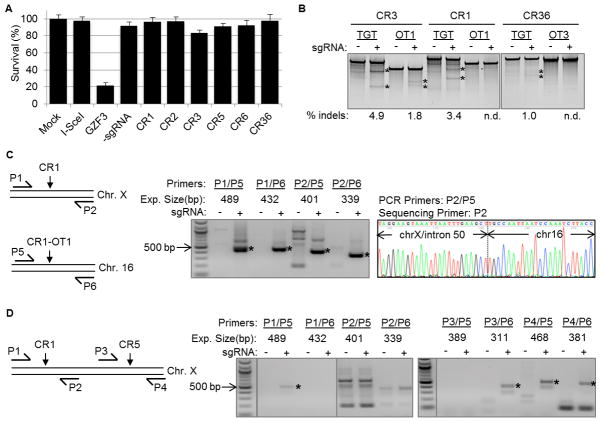
The CRISPR/Cas9 genome-editing platform is a promising technology to correct the genetic basis of hereditary diseases. The versatility, efficiency and multiplexing capabilities of the CRISPR/Cas9 system enable a variety of otherwise challenging gene correction strategies. Here, we use the CRISPR/Cas9 system to restore the expression of the dystrophin gene in cells carrying dystrophin mutations that cause Duchenne muscular dystrophy (DMD). We design single or multiplexed sgRNAs to restore the dystrophin reading frame by targeting the mutational hotspot at exons 45-55 and introducing shifts within exons or deleting one or more exons. Following gene editing in DMD patient myoblasts, dystrophin expression is restored in vitro. Human dystrophin is also detected in vivo after transplantation of genetically corrected patient cells into immunodeficient mice. Importantly, the unique multiplex gene-editing capabilities of the CRISPR/Cas9 system facilitate the generation of a single large deletion that can correct up to 62% of DMD mutations.
Conflict of interest statement
Figures

Comment in
-
Neuromuscular disease: CRISPR/Cas9 gene-editing platform corrects mutations associated with Duchenne muscular dystrophy.Nat Rev Neurol. 2015 Apr;11(4):184. doi: 10.1038/nrneurol.2015.37. Epub 2015 Mar 10. Nat Rev Neurol. 2015. PMID: 25752950 No abstract available.
-
Neuromuscular disease: Genome editing shows promise in an in vivo model of Duchenne muscular dystrophy.Nat Rev Neurol. 2016 Feb;12(2):63. doi: 10.1038/nrneurol.2016.4. Epub 2016 Jan 18. Nat Rev Neurol. 2016. PMID: 26782331 No abstract available.
References
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
