

The cadmium-tolerant pea (Pisum sativum L.) mutant SGECdt is more sensitive to mercury: assessing plant water relations
- PMID: 25694548
- PMCID: PMC4986718
- DOI: 10.1093/jxb/eru536
The cadmium-tolerant pea (Pisum sativum L.) mutant SGECdt is more sensitive to mercury: assessing plant water relations
Abstract
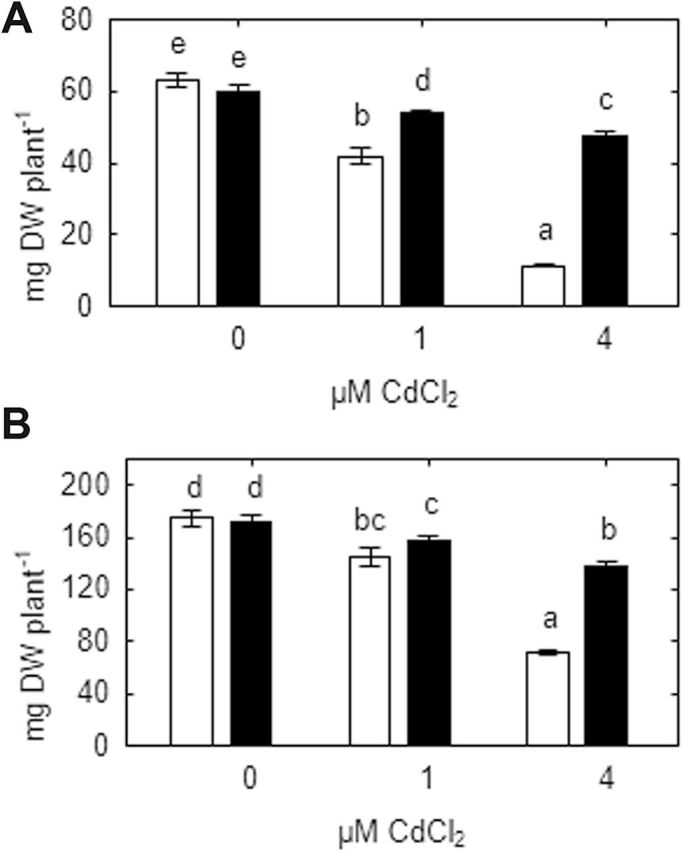
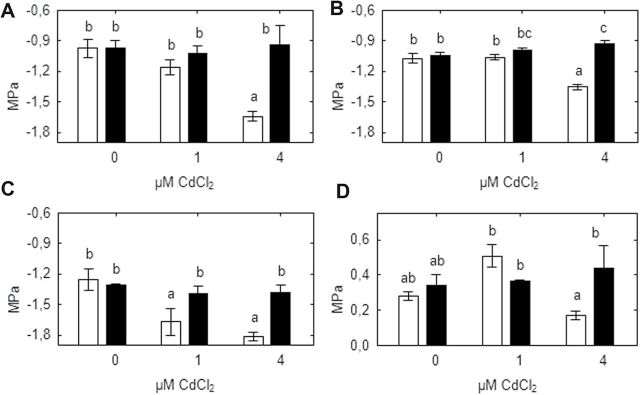
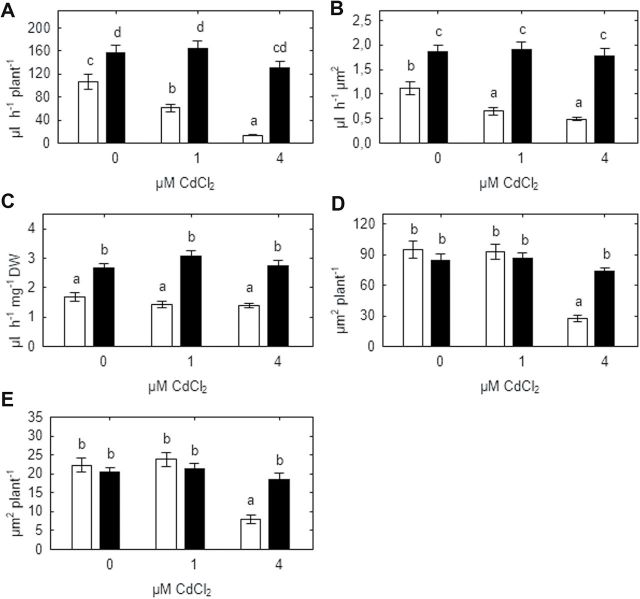
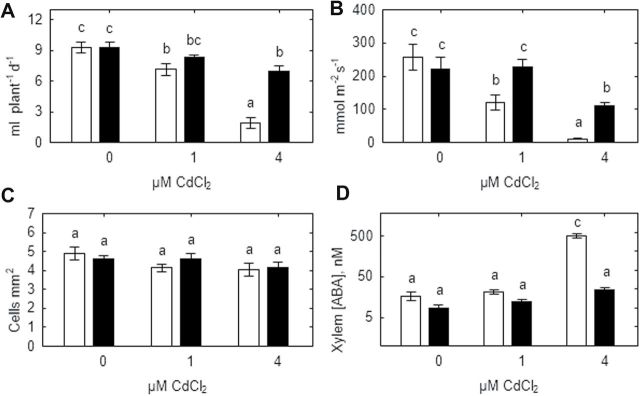
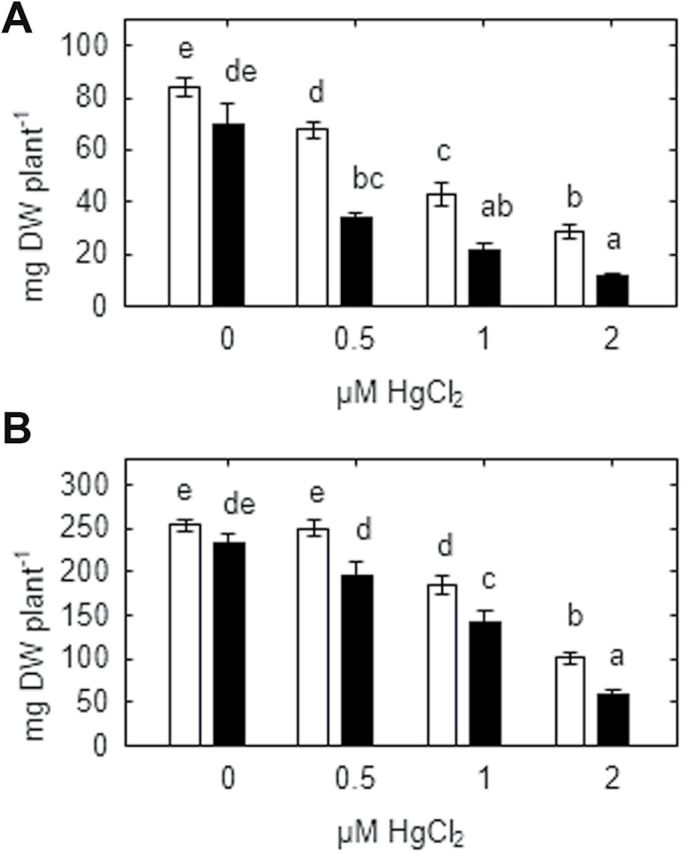
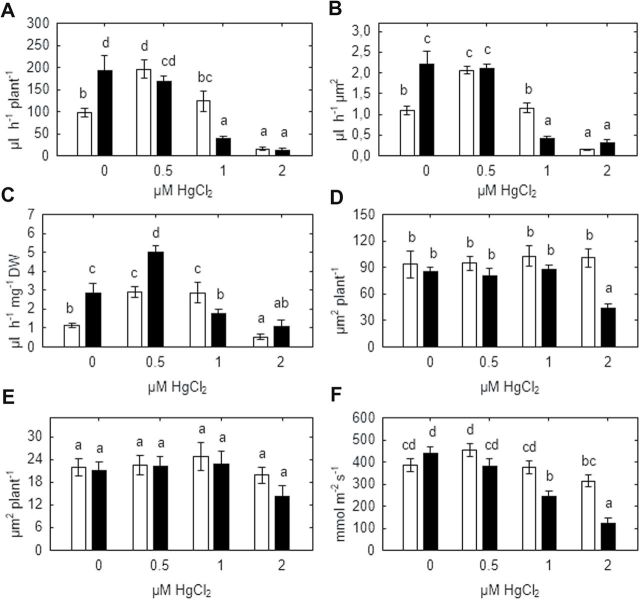
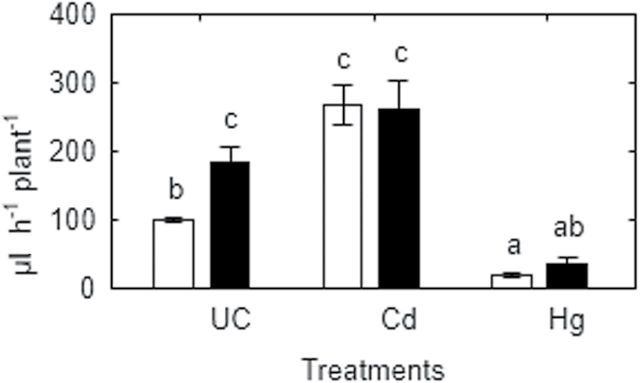
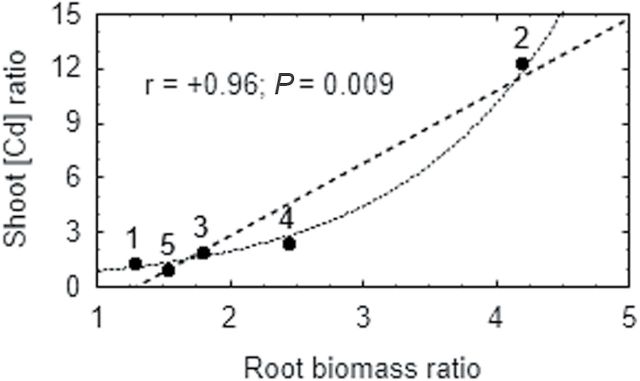
Heavy metals have multiple effects on plant growth and physiology, including perturbation of plant water status. These effects were assessed by exposing the unique Cd-tolerant and Cd-accumulating pea (Pisum sativum L.) mutant SGECd(t) and its wild-type (WT) line SGE to either cadmium (1, 4 μM CdCl2) or mercury (0.5, 1, 2 μM HgCl2) in hydroponic culture for 12 days. When exposed to Cd, SGECd(t) accumulated more Cd in roots, xylem sap, and shoot, and had considerably more biomass than WT plants. WT plants lost circa 0.2 MPa turgor when grown in 4 μM CdCl2, despite massive decreases in whole-plant transpiration rate and stomatal conductance. In contrast, root Hg accumulation was similar in both genotypes, but WT plants accumulated more Hg in leaves and had a higher stomatal conductance, and root and shoot biomass compared with SGECd(t). Shoot excision resulted in greater root-pressure induced xylem exudation of SGECd(t) in the absence of Cd or Hg and following Cd exposure, whereas the opposite response or no genotypic differences occurred following Hg exposure. Exposing plants that had not been treated with metal to 50 μM CdCl2 for 1h increased root xylem exudation of WT, whereas 50 μM HgCl2 inhibited and eliminated genotypic differences in root xylem exudation, suggesting differences between WT and SGECd(t) plants in aquaporin function. Thus, root water transport might be involved in mechanisms of increased tolerance and accumulation of Cd in the SGECd(t) mutant. However, the lack of cross-tolerance to Cd and Hg stress in the mutant indicates metal-specific mechanisms related to plant adaptation.
Keywords: Aquaporin; cadmium; drought; mercury; pea; root sap flow; water deficit..
© The Author 2015. Published by Oxford University Press on behalf of the Society for Experimental Biology. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures

References
-
- Aroca R., Porcel R., Ruiz-Lozano J.M. 2012. Regulation of root water uptake under abiotic stress conditions. Journal of Experimental Botany 63, 43–57. - PubMed
-
- Aroca R, Tognoni F, Irigoyen JJ, Sanchez-Diaz M, Pardossi A. 2001. Different root low temperature response of two maize genotypes differing in chilling sensitivity. Plant Physiology and Biochemistry 39, 1067–1073.
-
- Barceló J, Vázquez MD, Poshenrieder C. 1988. Cd induced structural and ultrastructural changes in the vascular system of bush bean stems. Botanica Acta 101, 254–261.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
