

Inducible HSP70 is critical in preventing the aggregation and enhancing the processing of PMP22
- PMID: 25694550
- PMCID: PMC4342366
- DOI: 10.1177/1759091415569909
Inducible HSP70 is critical in preventing the aggregation and enhancing the processing of PMP22
Abstract
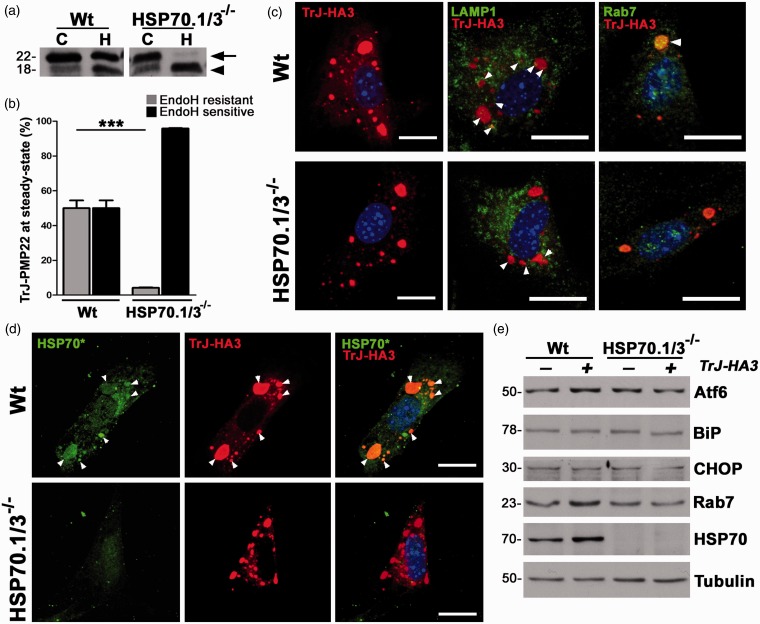
Chaperones, also called heat shock proteins (HSPs), transiently interact with proteins to aid their folding, trafficking, and degradation, thereby directly influencing the transport of newly synthesized molecules. Induction of chaperones provides a potential therapeutic approach for protein misfolding disorders, such as peripheral myelin protein 22 (PMP22)-associated peripheral neuropathies. Cytosolic aggregates of PMP22, linked with a demyelinating Schwann cell phenotype, result in suppression of proteasome activity and activation of proteostatic mechanisms, including the heat shock pathway. Although the beneficial effects of chaperones in preventing the aggregation and improving the trafficking of PMP22 have been repeatedly observed, the requirement for HSP70 in events remains elusive. In this study, we show that activation of the chaperone pathway in fibroblasts from PMP22 duplication-associated Charcot-Marie-Tooth disease type 1A patient with an FDA-approved small molecule increases HSP70 expression and attenuates proteasome dysfunction. Using cells from an HSP70.1/3(-/-) (inducible HSP70) mouse model, we demonstrate that under proteotoxic stress, this chaperone is critical in preventing the aggregation of PMP22, and this effect is aided by macroautophagy. When examined at steady-state, HSP70 appears to play a minor role in the trafficking of wild-type-PMP22, while it is crucial for preventing the buildup of the aggregation-prone Trembler-J-PMP22. HSP70 aids the processing of Trembler-J-PMP22 through the Golgi and its delivery to lysosomes via Rab7-positive vesicles. Together, these results demonstrate a key role for inducible HSP70 in aiding the processing and hindering the accumulation of misfolded PMP22, which in turn alleviates proteotoxicity within the cells.
Keywords: Charcot–Marie–Tooth disease; Trembler-J; chaperones; neuropathy; proteasome.
© The Author(s) 2015.
Figures






Similar articles
-
Biochemical characterization of protein quality control mechanisms during disease progression in the C22 mouse model of CMT1A.ASN Neuro. 2013 Dec 3;5(5):e00128. doi: 10.1042/AN20130024. ASN Neuro. 2013. PMID: 24175617 Free PMC article.
-
Impaired proteasome activity and accumulation of ubiquitinated substrates in a hereditary neuropathy model.J Neurochem. 2005 Mar;92(6):1531-41. doi: 10.1111/j.1471-4159.2004.02987.x. J Neurochem. 2005. PMID: 15748170
-
Emerging role for autophagy in the removal of aggresomes in Schwann cells.J Neurosci. 2003 Nov 19;23(33):10672-80. doi: 10.1523/JNEUROSCI.23-33-10672.2003. J Neurosci. 2003. PMID: 14627652 Free PMC article.
-
Regulation of myelin-specific gene expression. Relevance to CMT1.Ann N Y Acad Sci. 1999 Sep 14;883:91-108. Ann N Y Acad Sci. 1999. PMID: 10586235 Review.
-
The role of HSP70 and its co-chaperones in protein misfolding, aggregation and disease.Subcell Biochem. 2015;78:243-73. doi: 10.1007/978-3-319-11731-7_12. Subcell Biochem. 2015. PMID: 25487025 Review.
Cited by
-
Direct relationship between increased expression and mistrafficking of the Charcot-Marie-Tooth-associated protein PMP22.J Biol Chem. 2020 Aug 21;295(34):11963-11970. doi: 10.1074/jbc.AC120.014940. Epub 2020 Jul 9. J Biol Chem. 2020. PMID: 32647009 Free PMC article.
-
A brief review of recent Charcot-Marie-Tooth research and priorities.F1000Res. 2015 Feb 26;4:53. doi: 10.12688/f1000research.6160.1. eCollection 2015. F1000Res. 2015. PMID: 25901280 Free PMC article. Review.
-
Blocking mitochondrial calcium release in Schwann cells prevents demyelinating neuropathies.J Clin Invest. 2016 Mar 1;126(3):1023-38. doi: 10.1172/JCI84505. Epub 2016 Feb 15. J Clin Invest. 2016. Retraction in: J Clin Invest. 2017 Mar 1;127(3):1115. doi: 10.1172/JCI92100. PMID: 26878172 Free PMC article. Retracted.
-
Modulating Molecular Chaperones Improves Mitochondrial Bioenergetics and Decreases the Inflammatory Transcriptome in Diabetic Sensory Neurons.ACS Chem Neurosci. 2015 Sep 16;6(9):1637-48. doi: 10.1021/acschemneuro.5b00165. Epub 2015 Jul 22. ACS Chem Neurosci. 2015. PMID: 26161583 Free PMC article.
-
Targeting Heat Shock Protein 70 to Ameliorate c-Jun Expression and Improve Demyelinating Neuropathy.ACS Chem Neurosci. 2018 Feb 21;9(2):381-390. doi: 10.1021/acschemneuro.7b00377. Epub 2017 Nov 9. ACS Chem Neurosci. 2018. PMID: 29120605 Free PMC article.
References
-
- Baechner D., Liehr T., Hameister H., Altenberger H., Grehl H., Suter U., Rautenstrauss B. (1995) Widespread expression of the peripheral myelin protein-22 gene (PMP22) in neural and non-neural tissues during murine development. Journal of Neuroscience Research 42: 733–741. - PubMed
-
- Bennett E. J., Bence N. F., Jayakumar R., Kopito R. R. (2005) Global impairment of the ubiquitin-proteasome system by nuclear or cytoplasmic protein aggregates precedes inclusion body formation. Molecular Cell 17: 351–365. - PubMed
-
- Bobkova N. V., Garbuz D. G., Nesterova I., Medvinskaya N., Samokhin A., Alexandrova I., Evgen'ev M. (2014) Therapeutic effect of exogenous hsp70 in mouse models of Alzheimer's disease. Journal of Alzheimer’s Disease 38: 425–435. - PubMed
-
- Dantuma N. P., Lindsten K., Glas R., Jellne M., Masucci M. G. (2000) Short-lived green fluorescent proteins for quantifying ubiquitin/proteasome-dependent proteolysis in living cells. Nature Biotechnology 18: 538–543. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
