

Immunoprevention of chemical carcinogenesis through early recognition of oncogene mutations
- PMID: 25694611
- PMCID: PMC4355080
- DOI: 10.4049/jimmunol.1402125
Immunoprevention of chemical carcinogenesis through early recognition of oncogene mutations
Abstract
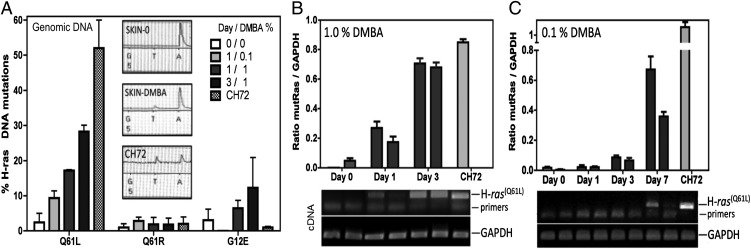
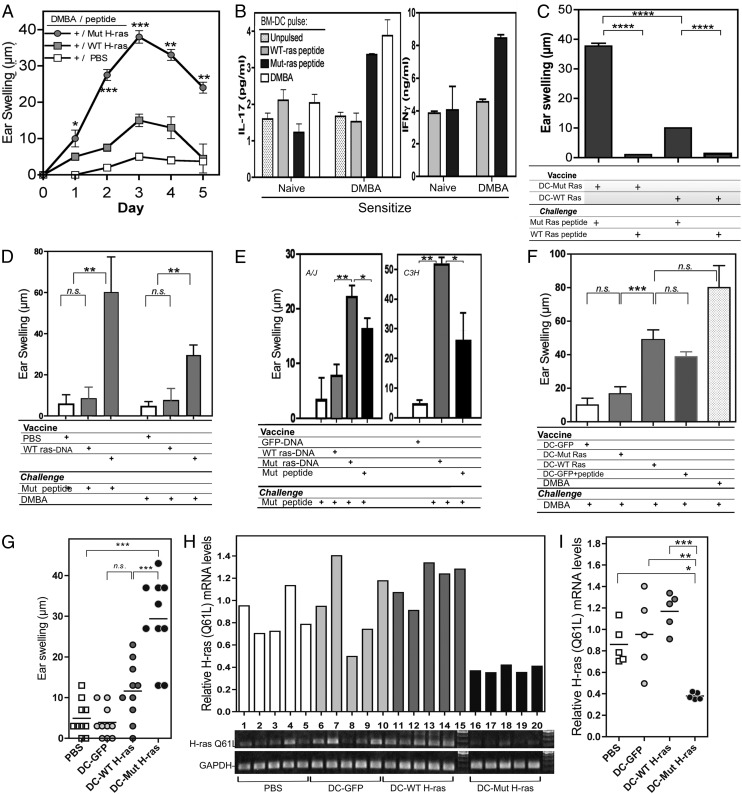
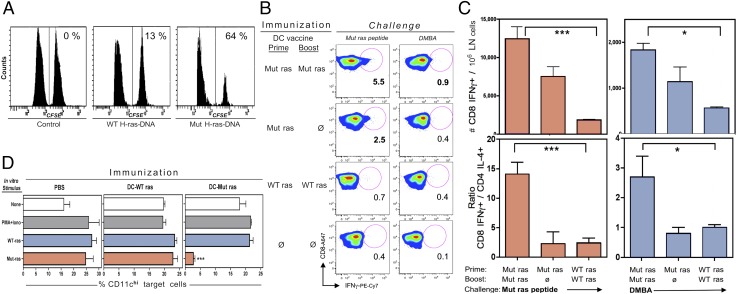
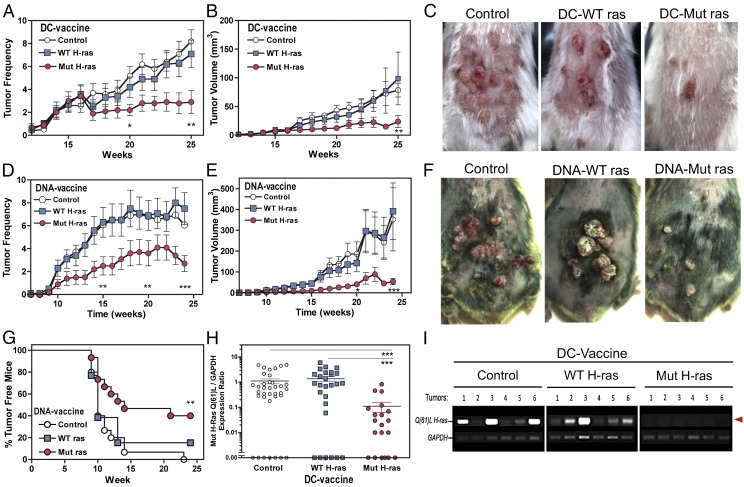
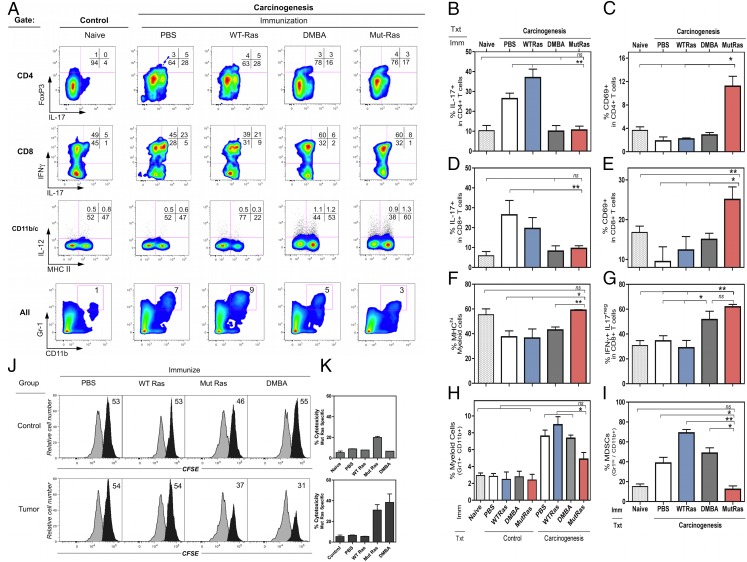
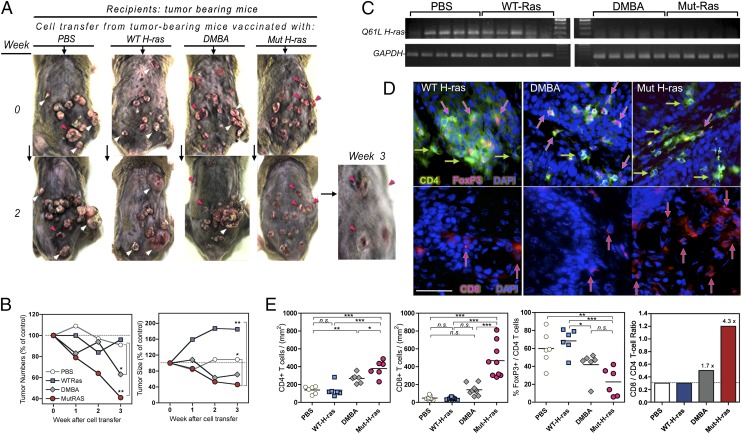
Prevention of tumors induced by environmental carcinogens has not been achieved. Skin tumors produced by polyaromatic hydrocarbons, such as 7,12-dimethylbenz(a)anthracene (DMBA), often harbor an H-ras point mutation, suggesting that it is a poor target for early immunosurveillance. The application of pyrosequencing and allele-specific PCR techniques established that mutations in the genome and expression of the Mut H-ras gene could be detected as early as 1 d after DMBA application. Further, DMBA sensitization raised Mut H-ras epitope-specific CTLs capable of eliminating Mut H-ras(+) preneoplastic skin cells, demonstrating that immunosurveillance is normally induced but may be ineffective owing to insufficient effector pool size and/or immunosuppression. To test whether selective pre-expansion of CD8 T cells with specificity for the single Mut H-ras epitope was sufficient for tumor prevention, MHC class I epitope-focused lentivector-infected dendritic cell- and DNA-based vaccines were designed to bias toward CTL rather than regulatory T cell induction. Mut H-ras, but not wild-type H-ras, epitope-focused vaccination generated specific CTLs and inhibited DMBA-induced tumor initiation, growth, and progression in preventative and therapeutic settings. Transferred Mut H-ras-specific effectors induced rapid tumor regression, overcoming established tumor suppression in tumor-bearing mice. These studies support further evaluation of oncogenic mutations for their potential to act as early tumor-specific, immunogenic epitopes in expanding relevant immunosurveillance effectors to block tumor formation, rather than treating established tumors.
Copyright © 2015 by The American Association of Immunologists, Inc.
Figures
Similar articles
-
Both (+/-)syn- and (+/-)anti-7,12-dimethylbenz[a]anthracene-3,4-diol-1,2-epoxides initiate tumors in mouse skin that possess -CAA- to -CTA- mutations at Codon 61 of c-H-ras.Cancer Res. 2000 Oct 15;60(20):5688-95. Cancer Res. 2000. PMID: 11059761
-
Rapid induction of skin tumors in human but not mouse c-Ha-ras proto-oncogene transgenic mice by chemical carcinogenesis.Cancer Sci. 2006 Sep;97(9):842-7. doi: 10.1111/j.1349-7006.2006.00256.x. Cancer Sci. 2006. PMID: 16805855 Free PMC article.
-
Identification of overlapping epitopes in mutant ras oncogene peptides that activate CD4+ and CD8+ T cell responses.Eur J Immunol. 1996 Feb;26(2):435-43. doi: 10.1002/eji.1830260225. Eur J Immunol. 1996. PMID: 8617315
-
Mutant ras epitopes as targets for cancer vaccines.Semin Oncol. 1996 Feb;23(1):118-34. Semin Oncol. 1996. PMID: 8607022 Review.
-
Non-melanoma skin cancer in mouse and man.Arch Toxicol. 2013 May;87(5):783-98. doi: 10.1007/s00204-012-0998-9. Epub 2012 Dec 25. Arch Toxicol. 2013. PMID: 23266722 Review.
Cited by
-
Potentiation of Kras peptide cancer vaccine by avasimibe, a cholesterol modulator.EBioMedicine. 2019 Nov;49:72-81. doi: 10.1016/j.ebiom.2019.10.044. Epub 2019 Oct 31. EBioMedicine. 2019. PMID: 31680003 Free PMC article.
-
Targets and Strategies for Cancer Immunoprevention.Methods Mol Biol. 2022;2435:7-17. doi: 10.1007/978-1-0716-2014-4_2. Methods Mol Biol. 2022. PMID: 34993936
-
The Tumor Microenvironment in SCC: Mechanisms and Therapeutic Opportunities.Front Cell Dev Biol. 2021 Feb 9;9:636544. doi: 10.3389/fcell.2021.636544. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 33634137 Free PMC article. Review.
-
Pathophysiological Integration of Metabolic Reprogramming in Breast Cancer.Cancers (Basel). 2022 Jan 10;14(2):322. doi: 10.3390/cancers14020322. Cancers (Basel). 2022. PMID: 35053485 Free PMC article. Review.
-
Cancer immunoprevention: from mice to early clinical trials.BMC Immunol. 2018 Jun 15;19(1):16. doi: 10.1186/s12865-018-0253-0. BMC Immunol. 2018. PMID: 29902992 Free PMC article. Review.
References
-
- Burnet F. M. 1970. The concept of immunological surveillance. Prog. Exp. Tumor Res. 13: 1–27. - PubMed
-
- Everall J. D., Dowd P. M. 1978. Influence of environmental factors excluding ultra violet radiation on the incidence of skin cancer. Bull. Cancer 65: 241–247. - PubMed
-
- Hoffmann D., Rivenson A., Hecht S. S. 1996. The biological significance of tobacco-specific N-nitrosamines: smoking and adenocarcinoma of the lung. Crit. Rev. Toxicol. 26: 199–211. - PubMed
-
- Hecht S. S. 2002. Cigarette smoking and lung cancer: chemical mechanisms and approaches to prevention. Lancet Oncol. 3: 461–469. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
