

MKK3 mediates inflammatory response through modulation of mitochondrial function
- PMID: 25697779
- PMCID: PMC4441852
- DOI: 10.1016/j.freeradbiomed.2015.01.035
MKK3 mediates inflammatory response through modulation of mitochondrial function
Abstract
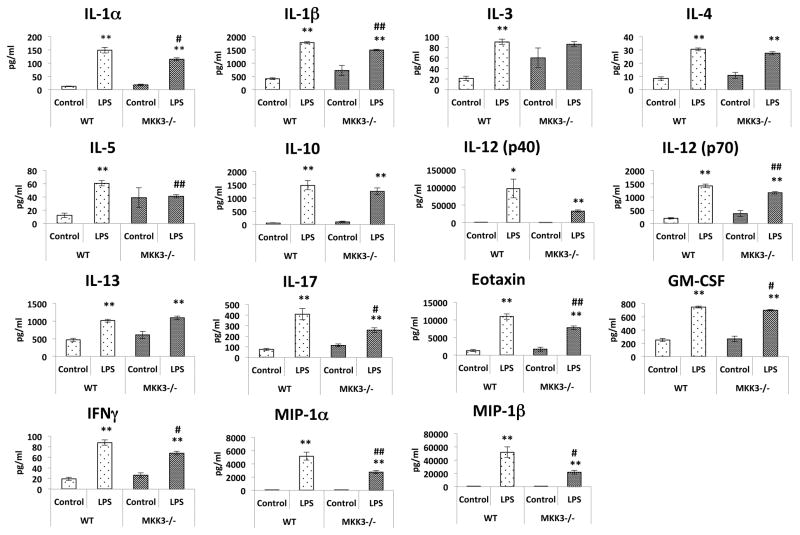
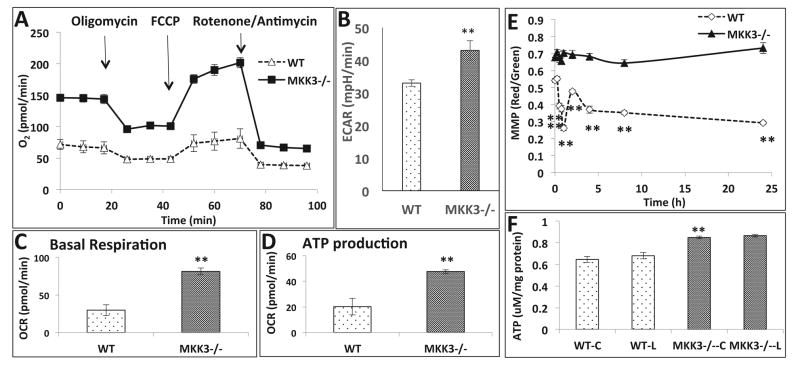
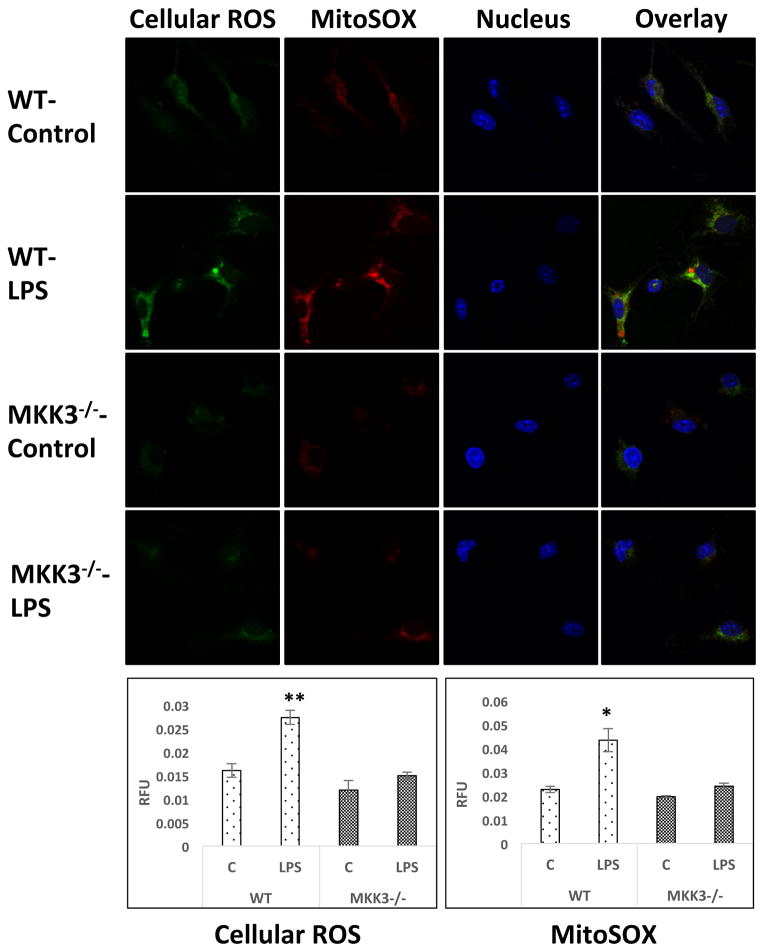
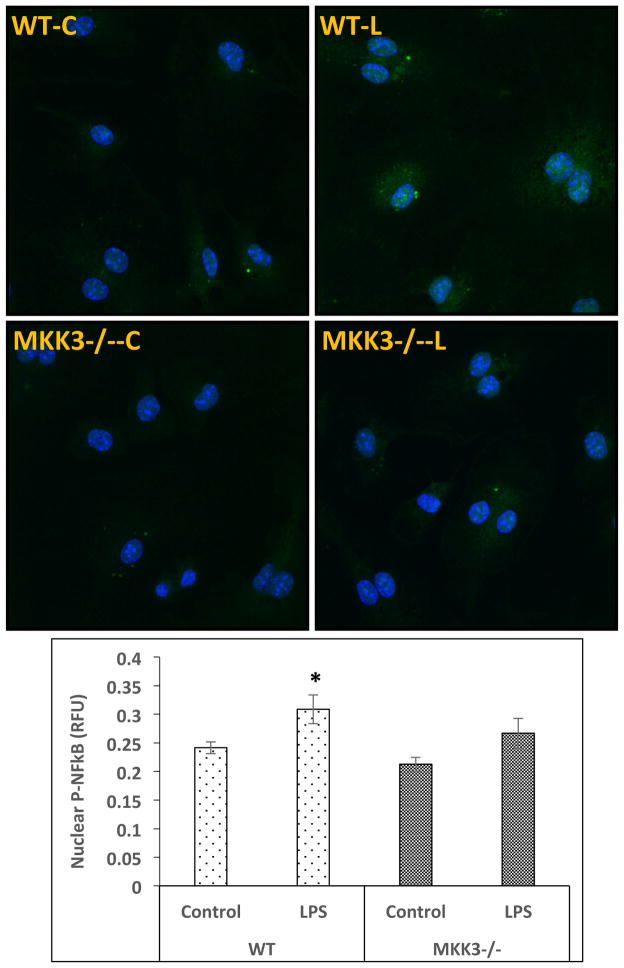
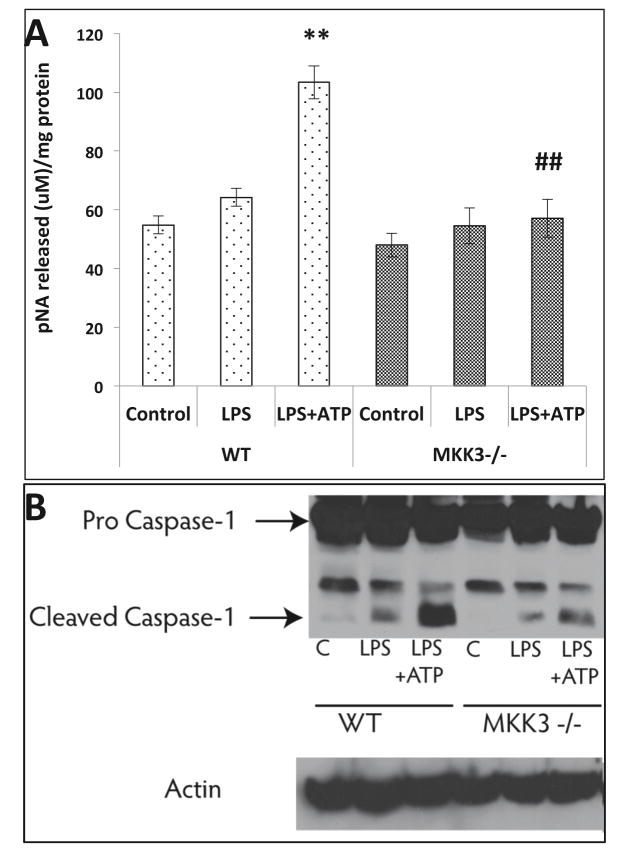
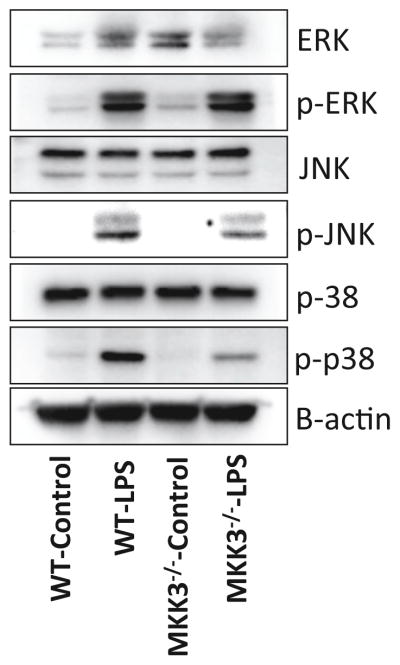
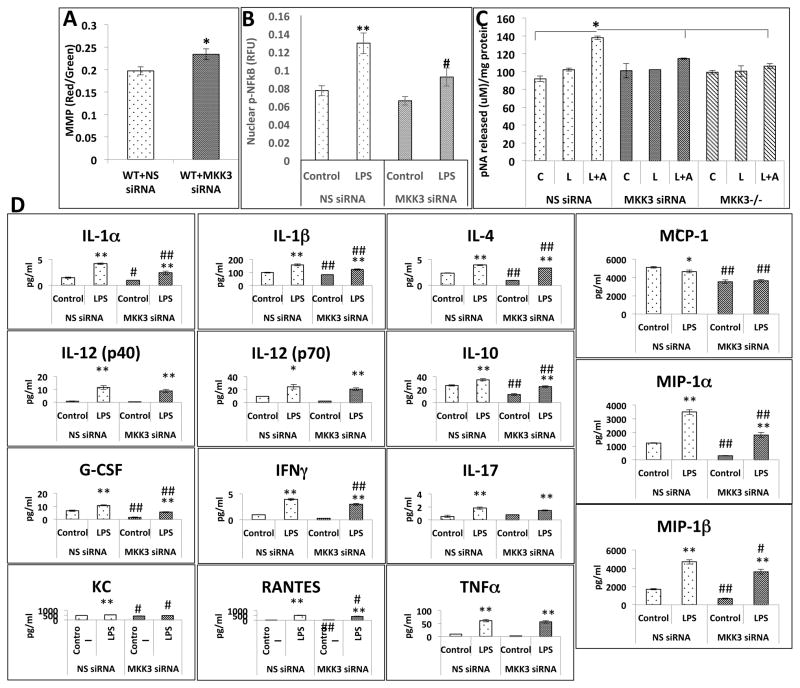
Mitochondria are increasingly recognized as drivers of inflammatory responses. MAP kinase kinase 3 (MKK3), a dual-specificity protein kinase, is activated in inflammation and in turn activates p38 MAP kinase signaling. Here we show that MKK3 influences mitochondrial function and acts as a critical mediator of inflammation. MKK3-deficient (MKK3(-/-)) mice and bone marrow-derived macrophages (BMDMs) secreted smaller amounts of cytokines than wild type (WT) after lipopolysaccharide (LPS) exposure. There was improved mitochondrial function, as measured by basal oxygen consumption rate, mitochondrial membrane potential, and ATP production, in MKK3(-/-) BMDMs. After LPS exposure, MKK3(-/-) BMDMs did not show a significant increase in cellular reactive oxygen species production or in mitochondrial superoxide compared to WT. Activation of two important inflammatory mediators, i.e., the nuclear translocation of NF-κB and caspase-1 activity (a key component of the inflammasome), was lower in MKK3(-/-) BMDMs. p38 and JNK activation was lower in MKK3(-/-) BMDMs compared to WT after exposure to LPS. Knockdown of MKK3 by siRNA in wild-type BMDMs improved mitochondrial membrane potential, reduced LPS-induced caspase-1 activation, and attenuated cytokine secretion. Our studies establish MKK3 as a regulator of mitochondrial function and inflammatory responses to LPS and suggest that MKK3 may be a therapeutic target in inflammatory disorders such as sepsis.
Keywords: Acute lung injury; Caspase 1; Free radicals; Inflammasome; Inflammation; Mitochondria; NF-κB; Oxidants; Sepsis.
Copyright © 2015 Elsevier Inc. All rights reserved.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the author(s).
Figures

References
-
- Huttemann M, Helling S, Sanderson TH, Sinkler C, Samavati L, Mahapatra G, Varughese A, Lu G, Liu J, Ramzan R, Vogt S, Grossman LI, Doan JW, Marcus K, Lee I. Regulation of mitochondrial respiration and apoptosis through cell signaling: cytochrome c oxidase and cytochrome c in ischemia/reperfusion injury and inflammation. Biochimica et biophysica acta. 2012;1817:598–609. - PMC - PubMed
-
- Debelec-Butuner B, Alapinar C, Varisli L, Erbaykent-Tepedelen B, Hamid SM, Gonen-Korkmaz C, Korkmaz KS. Inflammation-mediated abrogation of androgen signaling: an in vitro model of prostate cell inflammation. Molecular carcinogenesis. 2014;53:85–97. - PubMed
-
- Andrades ME, Ritter C, Dal-Pizzol F. The role of free radicals in sepsis development. Frontiers in bioscience. 2009;1:277–287. - PubMed
-
- Ryu MJ, Kim SJ, Kim YK, Choi MJ, Tadi S, Lee MH, Lee SE, Chung HK, Jung SB, Kim HJ, Jo YS, Kim KS, Lee SH, Kim JM, Kweon GR, Park KC, Lee JU, Kong YY, Lee CH, Chung J, Shong M. Crif1 deficiency reduces adipose OXPHOS capacity and triggers inflammation and insulin resistance in mice. PLoS genetics. 2013;9:e1003356. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
