

BiP and its nucleotide exchange factors Grp170 and Sil1: mechanisms of action and biological functions
- PMID: 25698114
- PMCID: PMC4356644
- DOI: 10.1016/j.jmb.2015.02.011
BiP and its nucleotide exchange factors Grp170 and Sil1: mechanisms of action and biological functions
Abstract
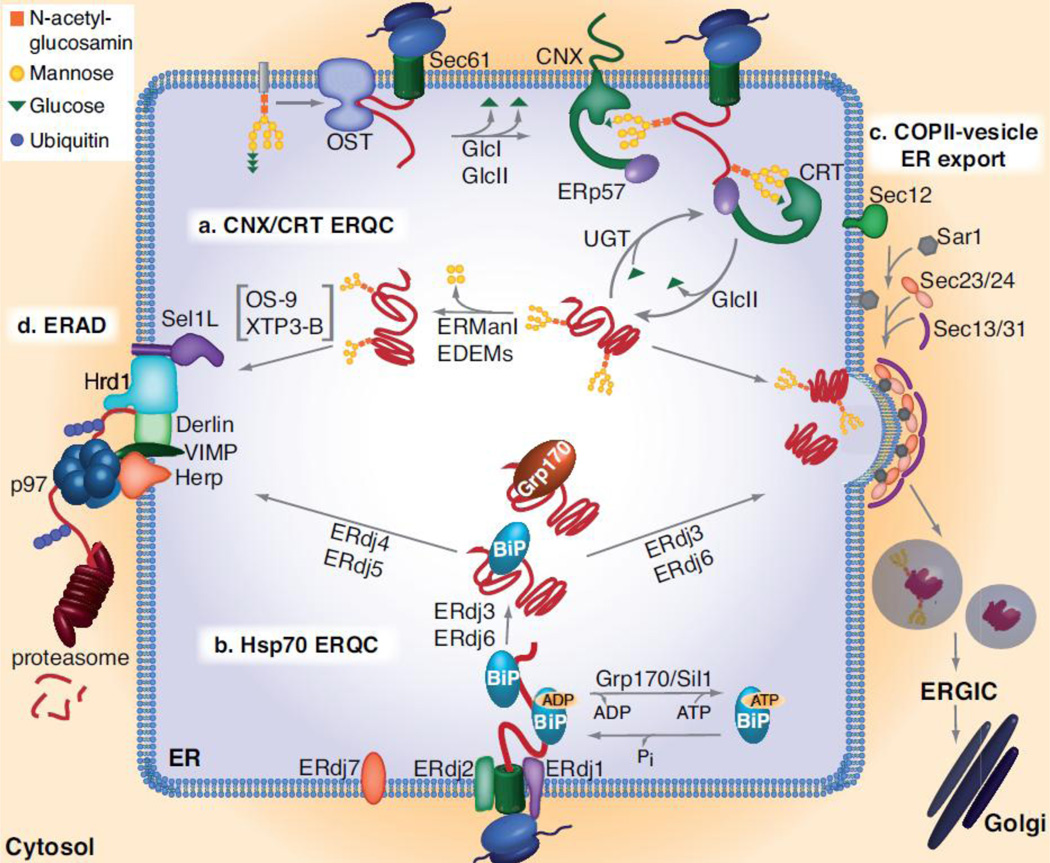
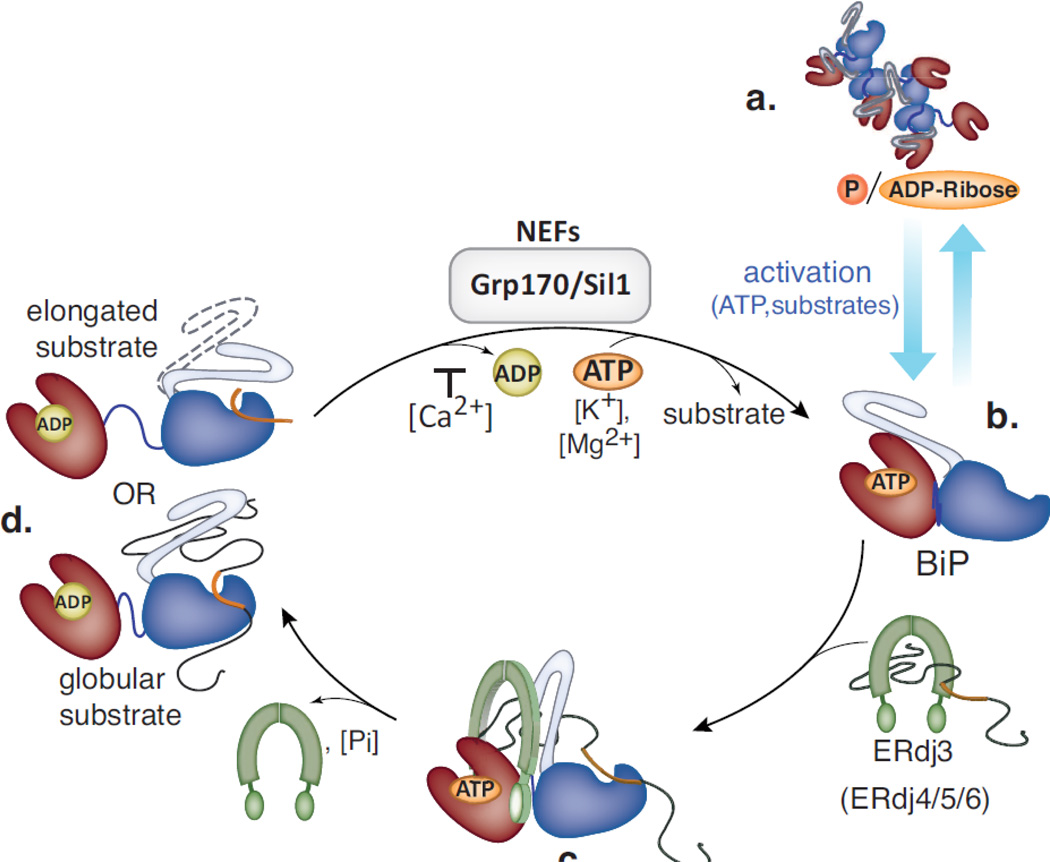
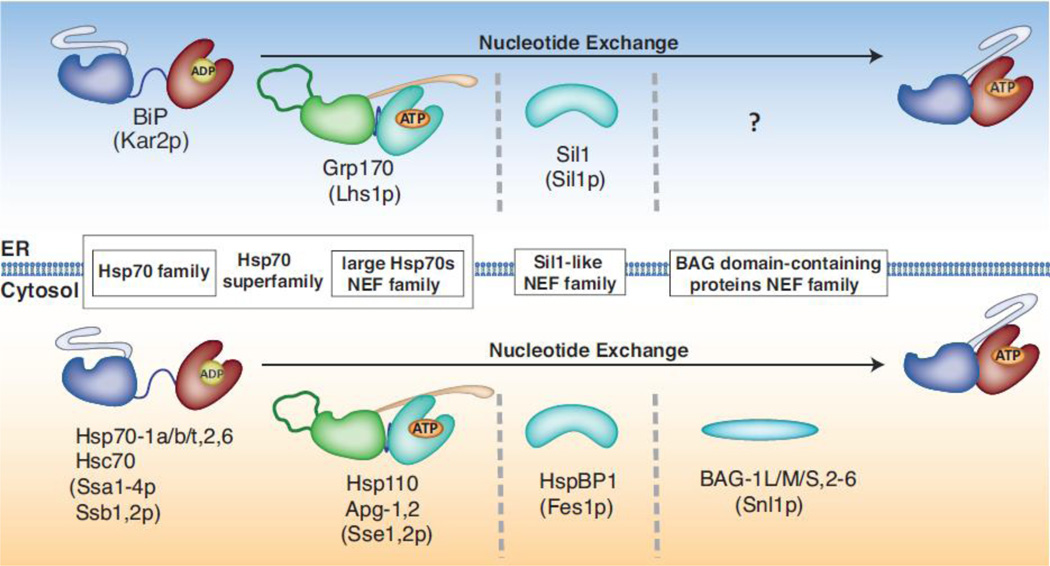
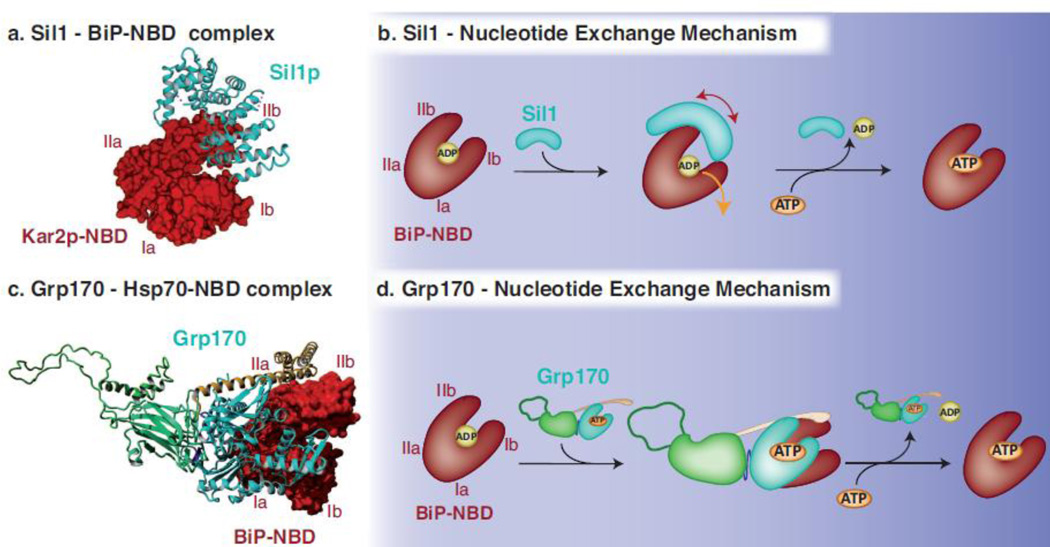
BiP (immunoglobulin heavy-chain binding protein) is the endoplasmic reticulum (ER) orthologue of the Hsp70 family of molecular chaperones and is intricately involved in most functions of this organelle through its interactions with a variety of substrates and regulatory proteins. Like all Hsp70 family members, the ability of BiP to bind and release unfolded proteins is tightly regulated by a cycle of ATP binding, hydrolysis, and nucleotide exchange. As a characteristic of the Hsp70 family, multiple DnaJ-like co-factors can target substrates to BiP and stimulate its ATPase activity to stabilize the binding of BiP to substrates. However, only in the past decade have nucleotide exchange factors for BiP been identified, which has shed light not only on the mechanism of BiP-assisted folding in the ER but also on Hsp70 family members that reside throughout the cell. We will review the current understanding of the ATPase cycle of BiP in the unique environment of the ER and how it is regulated by the nucleotide exchange factors, Grp170 (glucose-regulated protein of 170kDa) and Sil1, both of which perform unanticipated roles in various biological functions and disease states.
Keywords: BiP; Grp170; Sil1; nucleotide exchange factors; protein folding.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Molinari M, Helenius A. Chaperone selection during glycoprotein translocation into the endoplasmic reticulum. Science. 2000;288:331–333. - PubMed
-
- Melnyk A, Rieger H, Zimmermann R. Co-chaperones of the Mammalian Endoplasmic Reticulum. Subcell Biochem. 2015;78:179–200. - PubMed
-
- Chung KT, Shen Y, Hendershot LM. BAP, a mammalian BiP-associated protein, is a nucleotide exchange factor that regulates the ATPase activity of BiP. J Biol Chem. 2002;277:47557–47563. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
