

Cochlear injury and adaptive plasticity of the auditory cortex
- PMID: 25698966
- PMCID: PMC4318425
- DOI: 10.3389/fnagi.2015.00008
Cochlear injury and adaptive plasticity of the auditory cortex
Abstract
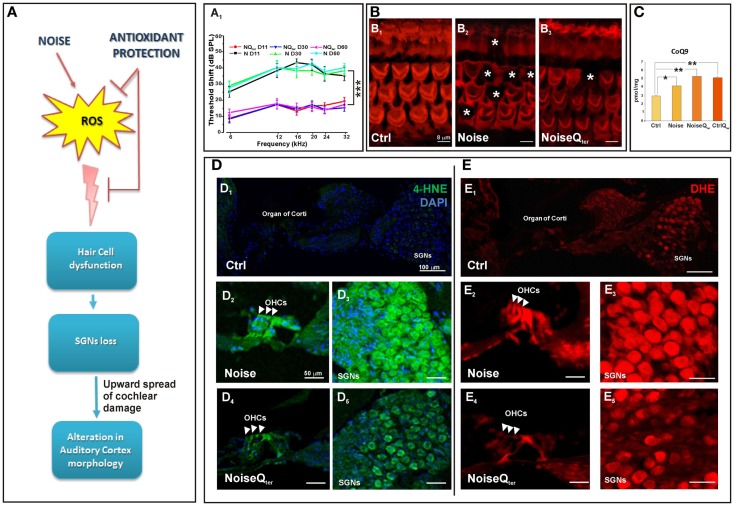
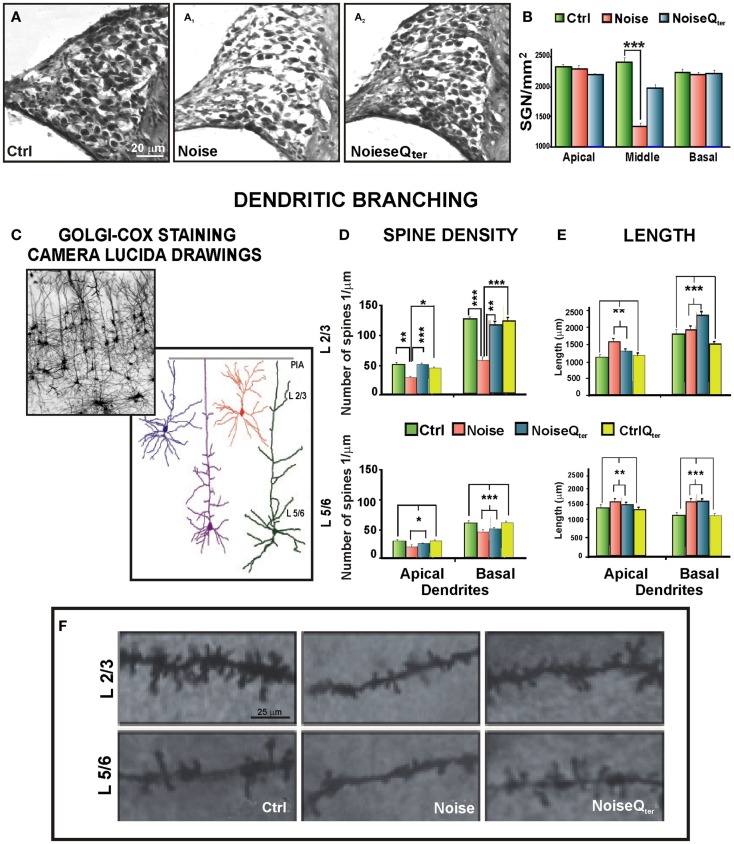
Growing evidence suggests that cochlear stressors as noise exposure and aging can induce homeostatic/maladaptive changes in the central auditory system from the brainstem to the cortex. Studies centered on such changes have revealed several mechanisms that operate in the context of sensory disruption after insult (noise trauma, drug-, or age-related injury). The oxidative stress is central to current theories of induced sensory-neural hearing loss and aging, and interventions to attenuate the hearing loss are based on antioxidant agent. The present review addresses the recent literature on the alterations in hair cells and spiral ganglion neurons due to noise-induced oxidative stress in the cochlea, as well on the impact of cochlear damage on the auditory cortex neurons. The emerging image emphasizes that noise-induced deafferentation and upward spread of cochlear damage is associated with the altered dendritic architecture of auditory pyramidal neurons. The cortical modifications may be reversed by treatment with antioxidants counteracting the cochlear redox imbalance. These findings open new therapeutic approaches to treat the functional consequences of the cortical reorganization following cochlear damage.
Keywords: auditory cortex; noise-induced hearing loss; oxidative stress; presbycusis; pyramidal neurons.
Figures
Similar articles
-
Noise-induced hearing loss (NIHL) as a target of oxidative stress-mediated damage: cochlear and cortical responses after an increase in antioxidant defense.J Neurosci. 2013 Feb 27;33(9):4011-23. doi: 10.1523/JNEUROSCI.2282-12.2013. J Neurosci. 2013. PMID: 23447610 Free PMC article.
-
Effects of lifetime noise exposure on the middle-age human auditory brainstem response, tinnitus and speech-in-noise intelligibility.Hear Res. 2018 Aug;365:36-48. doi: 10.1016/j.heares.2018.06.003. Epub 2018 Jun 12. Hear Res. 2018. PMID: 29913342
-
Candidate's thesis: enhancing intrinsic cochlear stress defenses to reduce noise-induced hearing loss.Laryngoscope. 2002 Sep;112(9):1515-32. doi: 10.1097/00005537-200209000-00001. Laryngoscope. 2002. PMID: 12352659
-
Insult-induced adaptive plasticity of the auditory system.Front Neurosci. 2014 May 23;8:110. doi: 10.3389/fnins.2014.00110. eCollection 2014. Front Neurosci. 2014. PMID: 24904256 Free PMC article. Review.
-
Pathology and mechanisms of cochlear aging.J Neurosci Res. 2020 Sep;98(9):1674-1684. doi: 10.1002/jnr.24439. Epub 2019 May 7. J Neurosci Res. 2020. PMID: 31066107 Free PMC article. Review.
Cited by
-
Cochlear Synaptopathy and Noise-Induced Hidden Hearing Loss.Neural Plast. 2016;2016:6143164. doi: 10.1155/2016/6143164. Epub 2016 Sep 21. Neural Plast. 2016. PMID: 27738526 Free PMC article. Review.
-
The evaluation of global cognitive and emotional status of older patients with chronic tinnitus.Brain Behav. 2021 Aug;11(8):e02074. doi: 10.1002/brb3.2074. Epub 2021 Jul 21. Brain Behav. 2021. PMID: 34288570 Free PMC article.
-
Tinnitus and Neuropsychological Dysfunction in the Elderly: A Systematic Review on Possible Links.J Clin Med. 2021 Apr 27;10(9):1881. doi: 10.3390/jcm10091881. J Clin Med. 2021. PMID: 33925344 Free PMC article. Review.
-
Physiological Evidence for Delayed Age-related Hearing Loss in Two Long-lived Rodent Species (Peromyscus leucopus and P. californicus).J Assoc Res Otolaryngol. 2022 Oct;23(5):617-631. doi: 10.1007/s10162-022-00860-4. Epub 2022 Jul 26. J Assoc Res Otolaryngol. 2022. PMID: 35882705 Free PMC article.
-
Effects of Aging and Adult-Onset Hearing Loss on Cortical Auditory Regions.Front Neurosci. 2016 May 11;10:199. doi: 10.3389/fnins.2016.00199. eCollection 2016. Front Neurosci. 2016. PMID: 27242405 Free PMC article. Review.
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources