

Comment
doi: 10.1016/j.molcel.2015.02.004.
TOPping off meiosis
Affiliations
- PMID: 25699706
- PMCID: PMC4352088
- DOI: 10.1016/j.molcel.2015.02.004
Item in Clipboard
Comment
TOPping off meiosis
Mol Cell.
.
Erratum in
- Mol Cell. 2015 Mar 19;57(6):1142
-
TOPping Off Meiosis.Mol Cell. 2015 Mar 19;57(6):1142. doi: 10.1016/j.molcel.2015.03.008. Epub 2015 Mar 19. Mol Cell. 2015. PMID: 29665396 No abstract available.
Abstract
Double-strand breaks (DSBs) threaten chromosome integrity. The most accurate repair of DSBs is by homologous recombination (HR), catalyzed by recombination proteins such as Rad51. Three papers in this issue of Molecular Cell (Fasching et al., 2015; Kaur et al., 2015; Tang et al., 2015) now reveal the role of three of these proteins in budding yeast: Sgs1 (BLM homolog), Top3 (TOPIIIα homolog), and Rmi1. They demonstrate several steps where all three proteins act together, and find additional functions of the Top3-Rmi1 subcomplex that are critical for the completion of meiosis.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
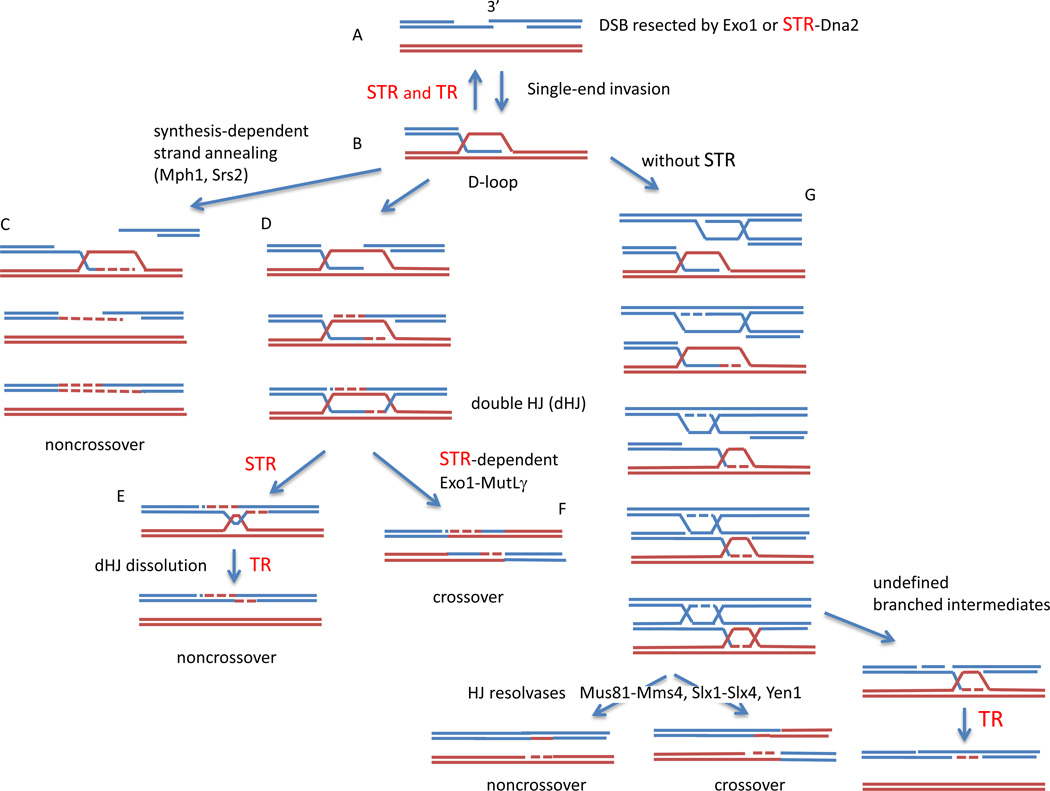
Roles of Sgs1, Top3 and Rmi1 in homologous recombination. A composite of steps in mitotic and meiotic recombination are shown, with the key steps requiring Sgs1-Top3-Rmi1 (STR) or Top3-Rmi1 alone (TR) shown in red. A. A double-strand break (DSB) is resected to yield 3’ ended single-strand DNA (ssDNA) tails either by the exonuclease Exo1 or by a helicase/endonuclease complex involving STR and Dna2. B. The ssDNA forms a nucleoprotein filament with Rad51 and engages in a search for homology, leading to single-end invasion and the formation of a D-loop. As shown by Fasching et al (2015) in this issue, D-loop formation can be reversed in two ways: by STR or by TR alone. C. The D-loop can be extended and the newly synthesized strand displaced, leading to DSB repair by synthesis-dependent strand annealing that yields noncrossover outcomes. D. The D-loop can be extended and result in an intermediate containing a fully ligated double Holliday junction (dHJ). The dHJ can be resolved into a noncrossover by dissolution, a process requiring STR to unwind and migrate the branched HJs and then TR to remove the remaining hemicatenane (E). Alternatively, the dHJ can be cleaved by Holliday junction resolvase. Here, the Exo1-MutLα complex that carries out this step in meiosis is shown (F). G. In the absence of STR, recombination in meiosis is greatly altered, with the appearance of multi-chromatid joint molecules (mcJMs) involving more than two of the 4 chromatids present at meiotic prophase (here three chromatids are illustrated). These mcJMs can be largely resolved by three structure-specific nucleases (Mus81-Mms4, Slx1-Slx4, or Yen1) into either crossover or noncrossover outcomes. However a minority of the intermediates formed without Sgs1 cannot be taken apart without the action of TR.
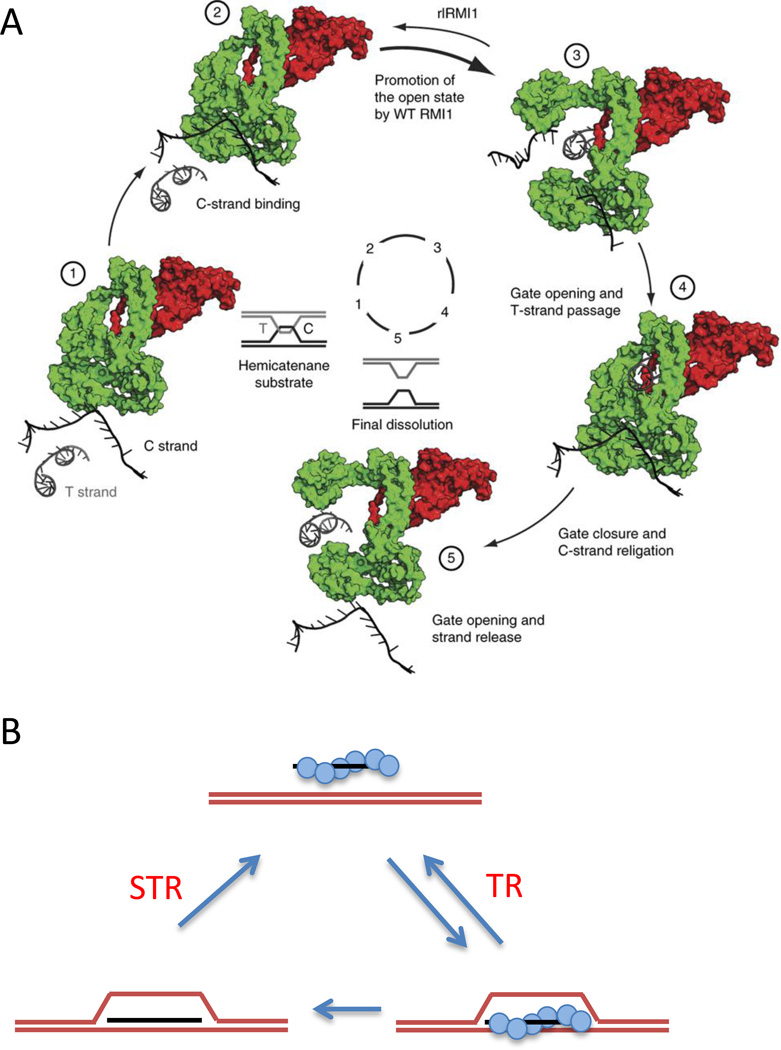
Roles of STR and TR in reversing the formation of D-loops. A. Mechanism of strand passage carried out by mammalian TOPIIIα-RMI1 as illustrated by (Bocquet et al., 2014). TopIIIα cleaves the C strand and undergoes a conformational change that allows the transfer strand (T) to pass through, after which the C strand is re-ligated and the gate closes, with release of the C strand. This action is stimulated by a loop of RMI1 that is part of the active site. Figure reused with permission from Bocquet et al., NSMB 2014 Figure 5. B. Rad51 (blue circles) coating single-stranded DNA (ssDNA) facilitates strand invasion and the formation of a D-loop in the presence of the ssDNA binding protein complex, RPA, and Rad54. When the D-loop is protein-free, Sgs1 alone, or STR, can take apart the D-loop, but Sgs1 alone cannot dismantle the protein-bound. This protein-bound form can be taken apart by the Top3-Rmi1 complex acting alone.
Comment on
-
Top3-Rmi1 DNA single-strand decatenase is integral to the formation and resolution of meiotic recombination intermediates.Mol Cell. 2015 Feb 19;57(4):583-594. doi: 10.1016/j.molcel.2015.01.020. Mol Cell. 2015. PMID: 25699707 Free PMC article.
-
Top3-Rmi1 dissolve Rad51-mediated D loops by a topoisomerase-based mechanism.Mol Cell. 2015 Feb 19;57(4):595-606. doi: 10.1016/j.molcel.2015.01.022. Mol Cell. 2015. PMID: 25699708 Free PMC article.
-
Pervasive and essential roles of the Top3-Rmi1 decatenase orchestrate recombination and facilitate chromosome segregation in meiosis.Mol Cell. 2015 Feb 19;57(4):607-621. doi: 10.1016/j.molcel.2015.01.021. Mol Cell. 2015. PMID: 25699709 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
