

Top3-Rmi1 dissolve Rad51-mediated D loops by a topoisomerase-based mechanism
- PMID: 25699708
- PMCID: PMC4338411
- DOI: 10.1016/j.molcel.2015.01.022
Top3-Rmi1 dissolve Rad51-mediated D loops by a topoisomerase-based mechanism
Abstract
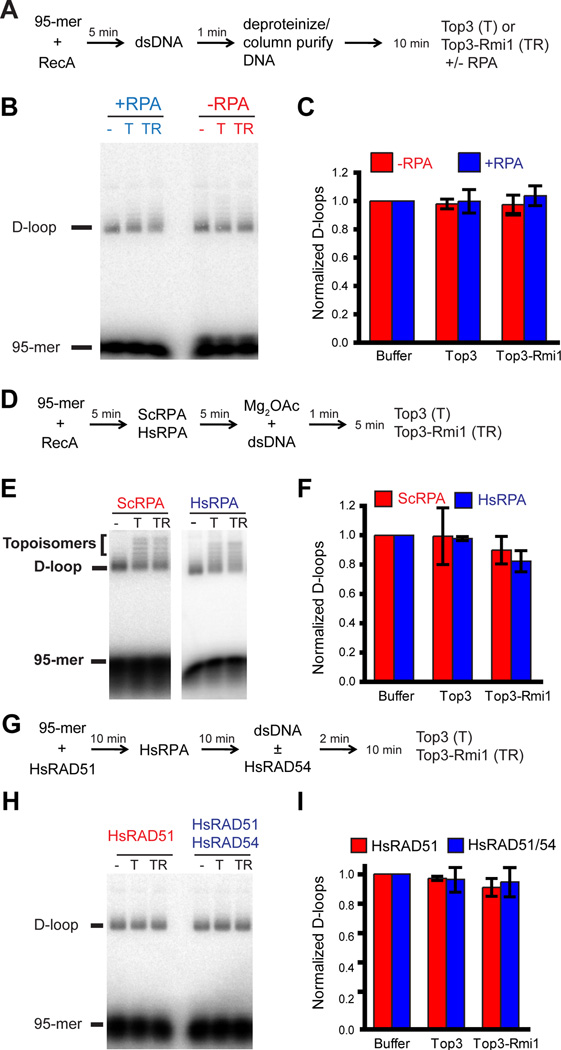
The displacement loop (D loop) is a DNA strand invasion product formed during homologous recombination. Disruption of nascent D loops prevents recombination, and during synthesis-dependent strand annealing (SDSA), disruption of D loops extended by DNA polymerase ensures a non-crossover outcome. The proteins implicated in D loop disruption are DNA motor proteins/helicases that act by moving DNA junctions. Here we report that D loops can also be disrupted by DNA topoisomerase 3 (Top3), and this disruption depends on Top3's catalytic activity. Yeast Top3 specifically disrupts D loops mediated by yeast Rad51/Rad54; protein-free D loops or D loop mediated by bacterial RecA protein or human RAD51/RAD54 resist dissolution. Also, the human Topoisomerase IIIa-RMI1-RMI2 complex is capable of dissolving D loops. Consistent with genetic data, we suggest that the extreme growth defect and hyper-recombination phenotype of Top3-deficient yeast cells is partially a result of unprocessed D loops.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures





Comment in
-
TOPping off meiosis.Mol Cell. 2015 Feb 19;57(4):577-581. doi: 10.1016/j.molcel.2015.02.004. Mol Cell. 2015. PMID: 25699706 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
- F32 GM083509/GM/NIGMS NIH HHS/United States
- R37 GM062653/GM/NIGMS NIH HHS/United States
- CA92276/CA/NCI NIH HHS/United States
- R01 GM062653/GM/NIGMS NIH HHS/United States
- R01 CA092276/CA/NCI NIH HHS/United States
- GM58015/GM/NIGMS NIH HHS/United States
- F32 GM83509/GM/NIGMS NIH HHS/United States
- R01 GM058015/GM/NIGMS NIH HHS/United States
- GM41347/GM/NIGMS NIH HHS/United States
- R01 CA154920/CA/NCI NIH HHS/United States
- R01 GM041347/GM/NIGMS NIH HHS/United States
- CA154920/CA/NCI NIH HHS/United States
- GM62653/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
