

Pervasive and essential roles of the Top3-Rmi1 decatenase orchestrate recombination and facilitate chromosome segregation in meiosis
- PMID: 25699709
- PMCID: PMC4791043
- DOI: 10.1016/j.molcel.2015.01.021
Pervasive and essential roles of the Top3-Rmi1 decatenase orchestrate recombination and facilitate chromosome segregation in meiosis
Abstract
The Bloom's helicase ortholog, Sgs1, plays central roles to coordinate the formation and resolution of joint molecule intermediates (JMs) during meiotic recombination in budding yeast. Sgs1 can associate with type-I topoisomerase Top3 and its accessory factor Rmi1 to form a conserved complex best known for its unique ability to decatenate double-Holliday junctions. Contrary to expectations, we show that the strand-passage activity of Top3-Rmi1 is required for all known functions of Sgs1 in meiotic recombination, including channeling JMs into physiological crossover and noncrossover pathways, and suppression of non-allelic recombination. We infer that Sgs1 always functions in the context of the Sgs1-Top3-Rmi1 complex to regulate meiotic recombination. In addition, we reveal a distinct late role for Top3-Rmi1 in resolving recombination-dependent chromosome entanglements to allow segregation at anaphase. Surprisingly, Sgs1 does not share this essential role of Top3-Rmi1. These data reveal an essential and pervasive role for the Top3-Rmi1 decatenase during meiosis.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
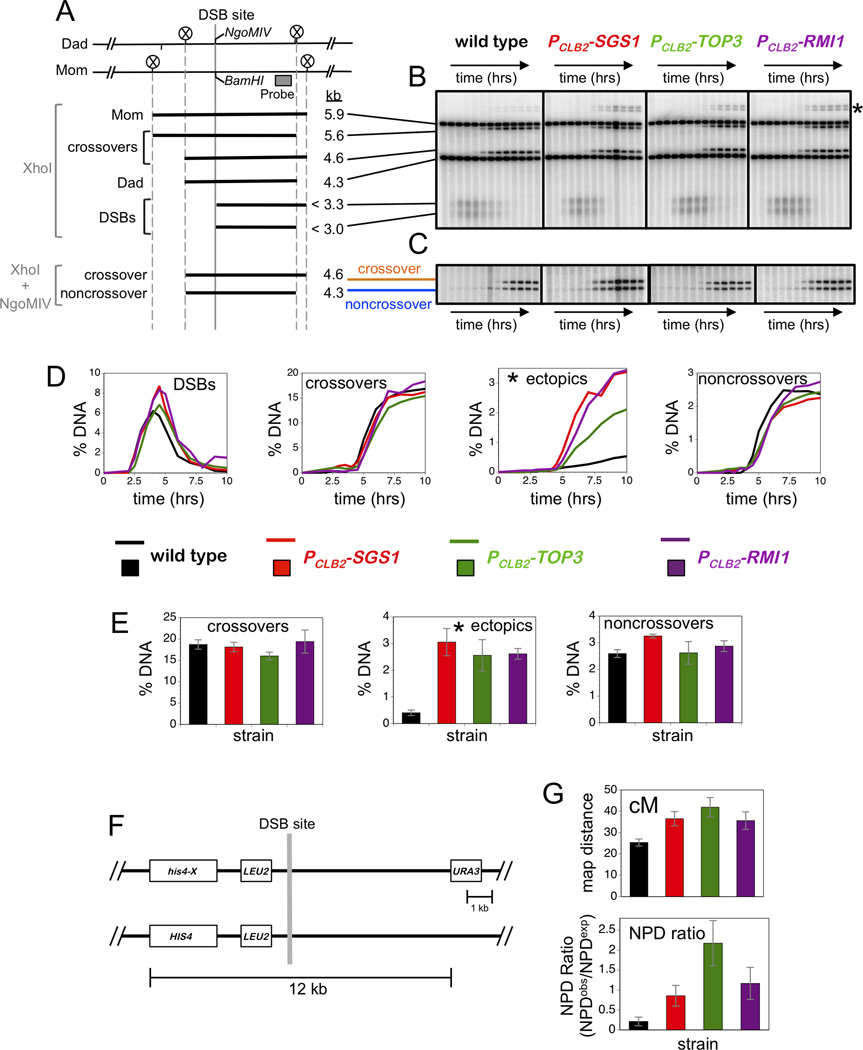
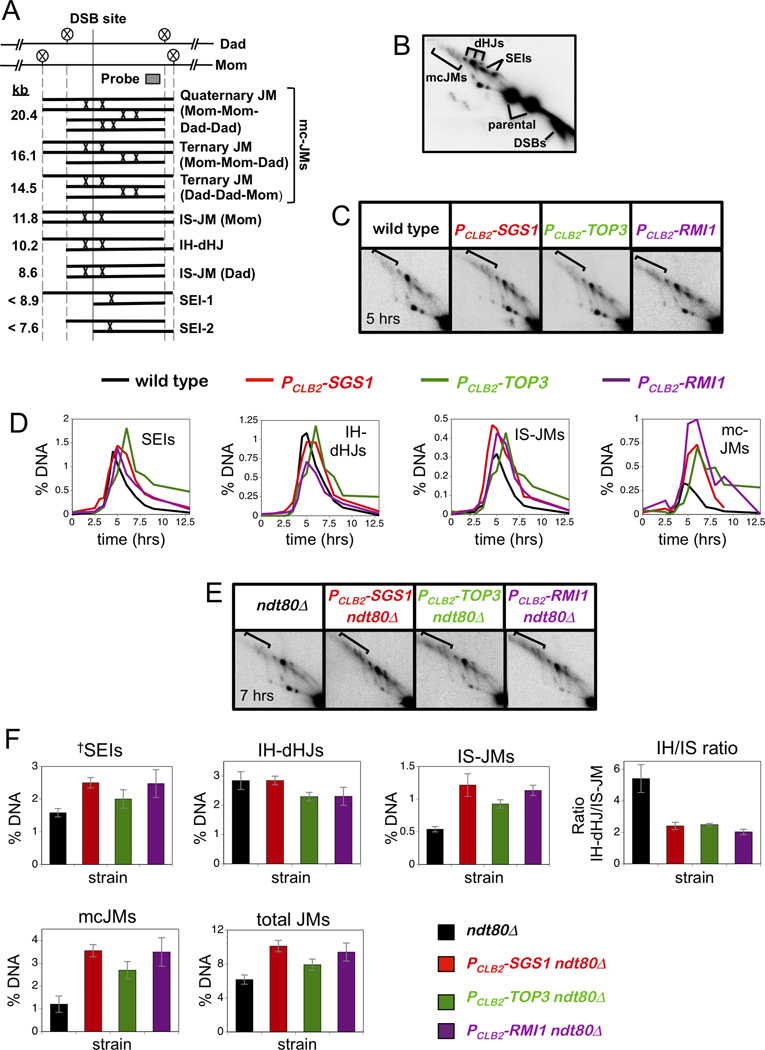
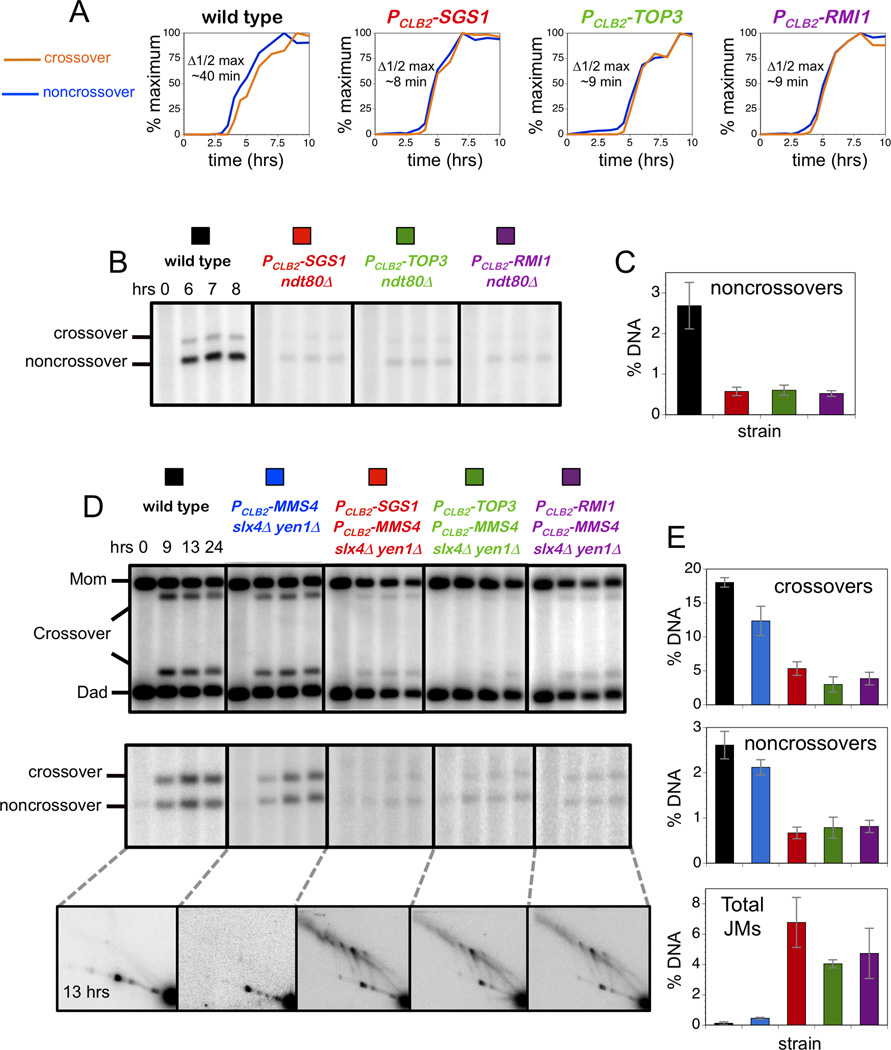
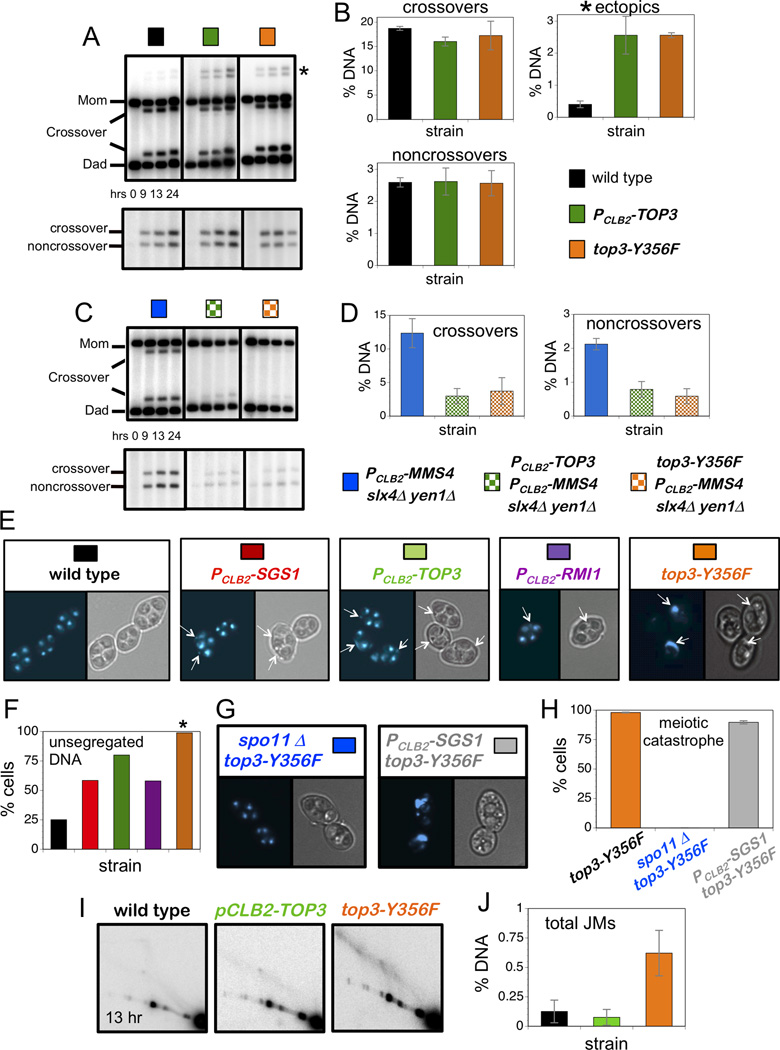
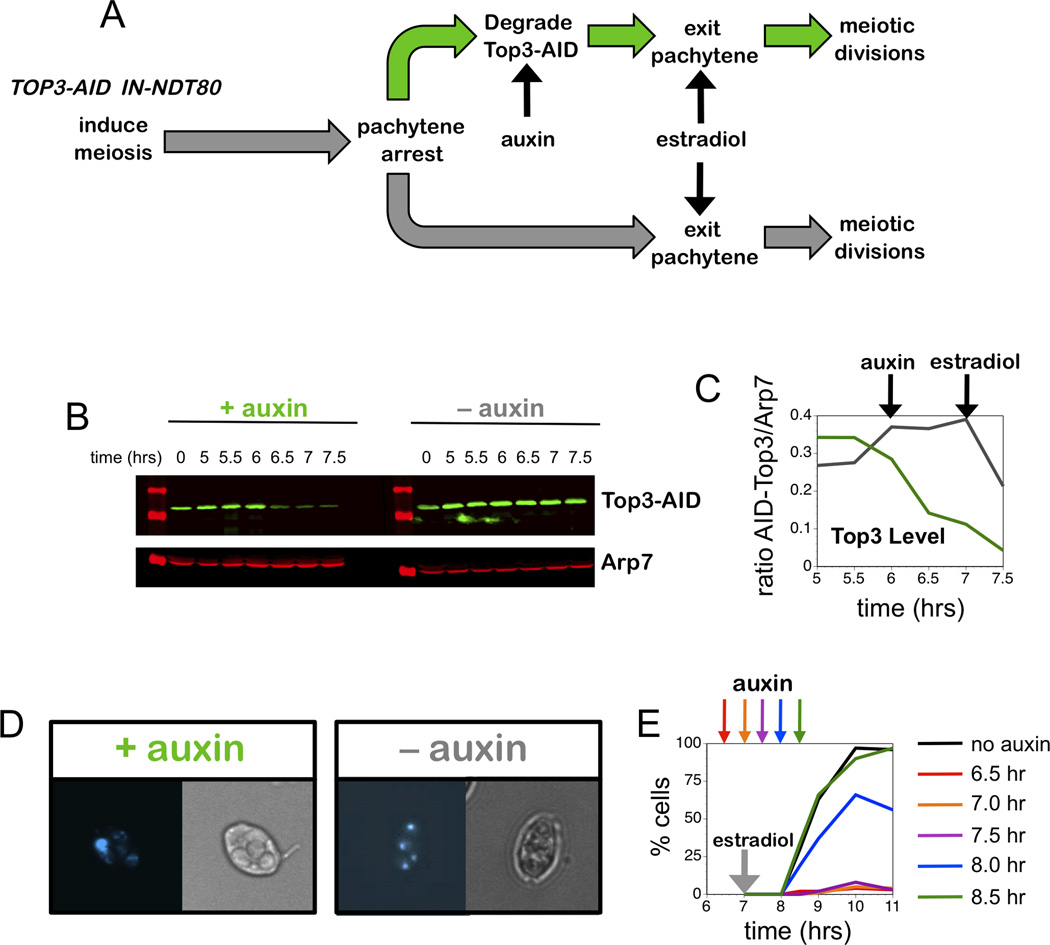
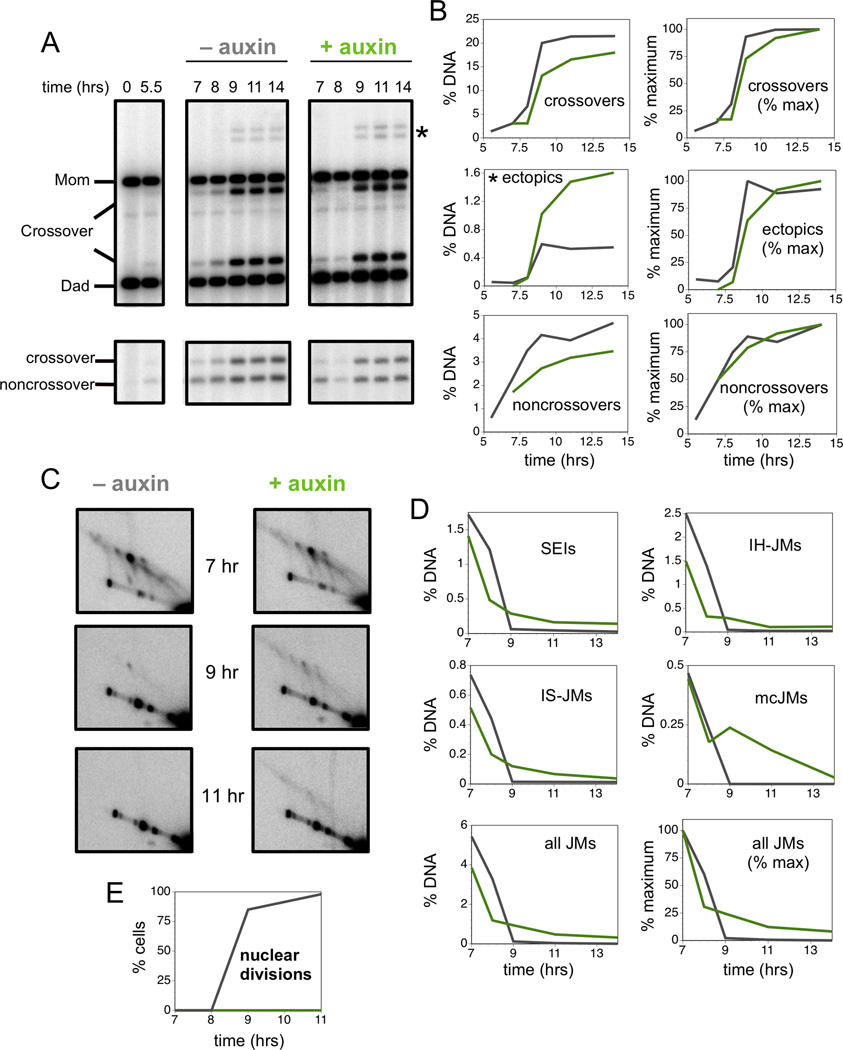
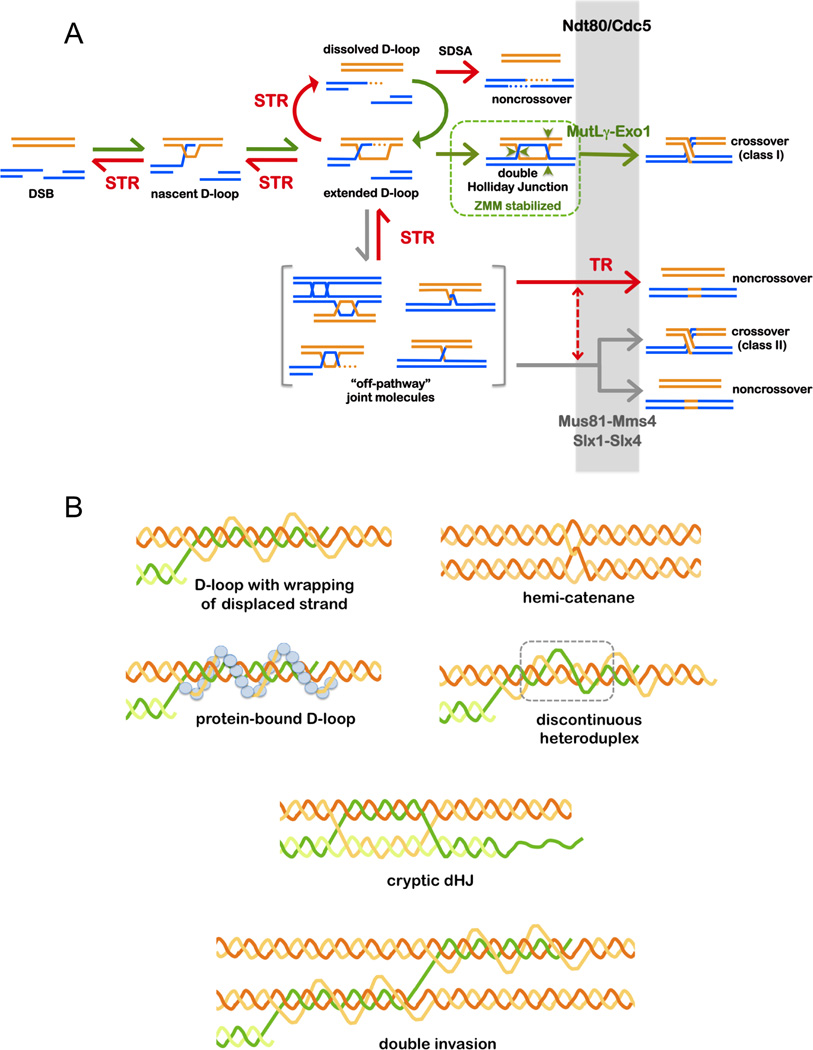
Comment in
-
TOPping off meiosis.Mol Cell. 2015 Feb 19;57(4):577-581. doi: 10.1016/j.molcel.2015.02.004. Mol Cell. 2015. PMID: 25699706 Free PMC article.
References
-
- Allers T, Lichten M. Differential timing and control of noncrossover and crossover recombination during meiosis. Cell. 2001;106:47–57. - PubMed
-
- Andersen SL, Sekelsky J. Meiotic versus mitotic recombination: two different routes for double-strand break repair: the different functions of meiotic versus mitotic DSB repair are reflected in different pathway usage and different outcomes. BioEssays : news and reviews in molecular, cellular and developmental biology. 2010;32:1058–1066. - PMC - PubMed
-
- Bell L, Byers B. Separation of branched from linear DNA by two-dimensional gel electrophoresis. Anal Biochem. 1983;130:527–535. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
