

The adult murine heart has a sparse, phagocytically active macrophage population that expands through monocyte recruitment and adopts an 'M2' phenotype in response to Th2 immunologic challenge
- PMID: 25700973
- PMCID: PMC4451497
- DOI: 10.1016/j.imbio.2015.01.013
The adult murine heart has a sparse, phagocytically active macrophage population that expands through monocyte recruitment and adopts an 'M2' phenotype in response to Th2 immunologic challenge
Abstract
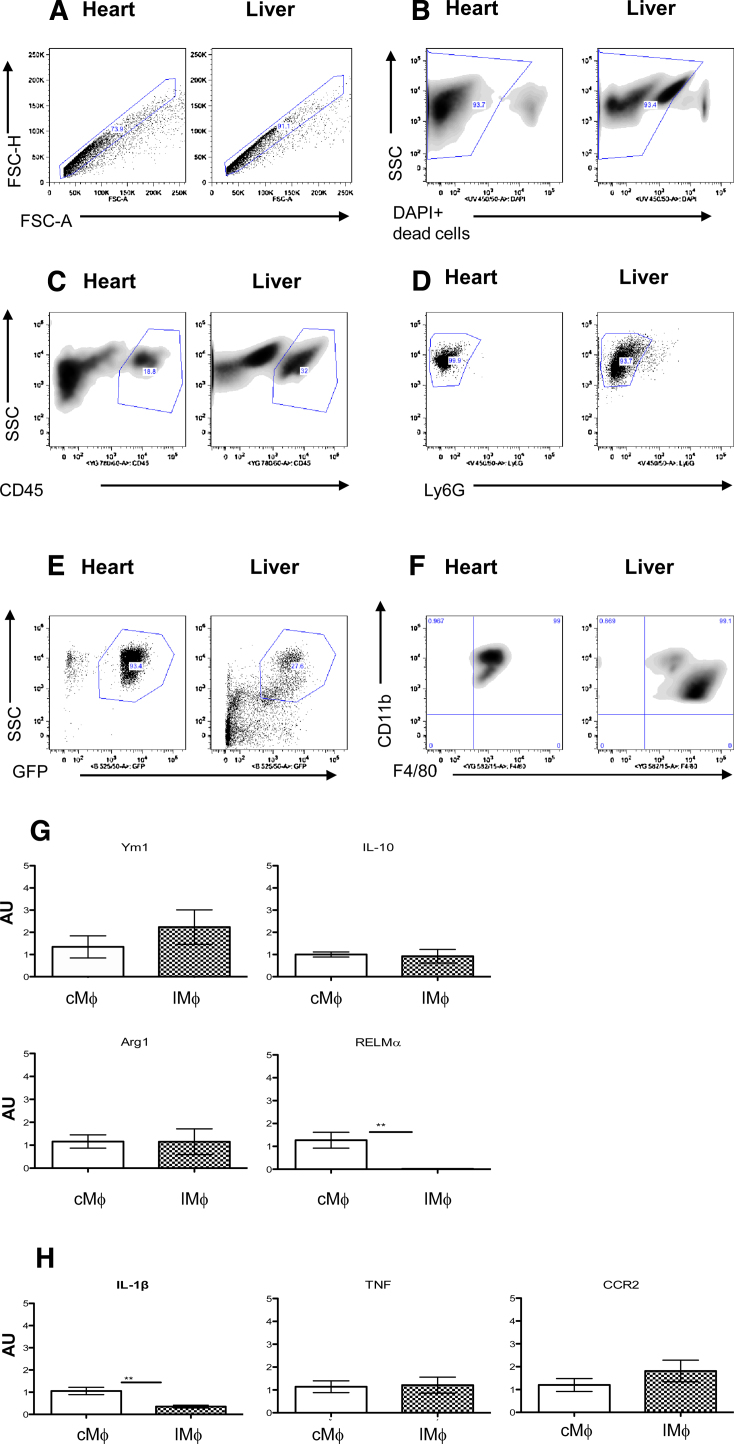
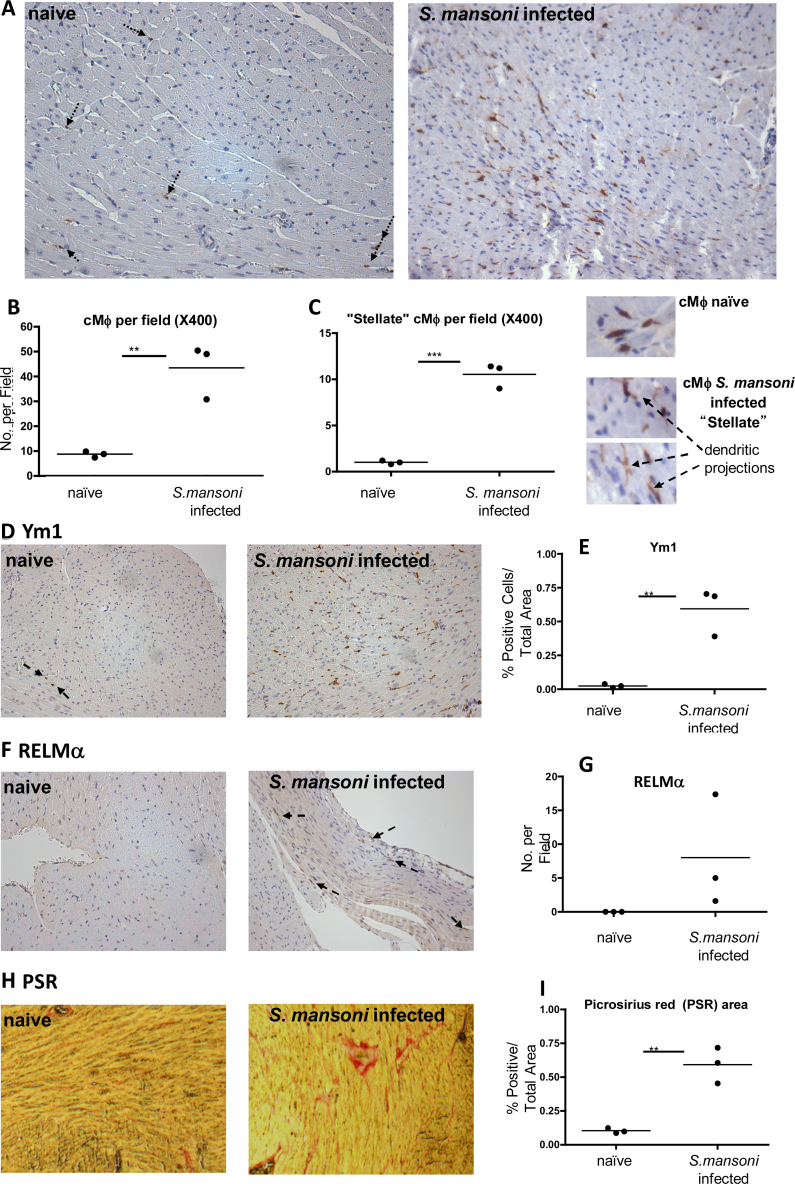
Tissue resident macrophages have vital homeostatic roles in many tissues but their roles are less well defined in the heart. The present study aimed to identify the density, polarisation status and distribution of macrophages in the healthy murine heart and to investigate their ability to respond to immune challenge. Histological analysis of hearts from CSF-1 receptor (csf1-GFP; MacGreen) and CX3CR1 (Cx3cr1(GFP/+)) reporter mice revealed a sparse population of GFP positive macrophages that were evenly distributed throughout the left and right ventricular free walls and septum. F4/80+CD11b+ cardiac macrophages, sorted from myocardial homogenates, were able to phagocytose fluorescent beads in vitro and expressed markers typical of both 'M1' (IL-1β, TNF and CCR2) and 'M2' activation (Ym1, Arg 1, RELMα and IL-10), suggesting no specific polarisation in healthy myocardium. Exposure to Th2 challenge by infection of mice with helminth parasites Schistosoma mansoni, or Heligmosomoides polygyrus, resulted in an increase in cardiac macrophage density, adoption of a stellate morphology and increased expression of Ym1, RELMα and CD206 (mannose receptor), indicative of 'M2' polarisation. This was dependent on recruitment of Ly6ChighCCR2+ monocytes and was accompanied by an increase in collagen content. In conclusion, in the healthy heart resident macrophages are relatively sparse and have a phagocytic role. Following Th2 challenge this population expands due to monocyte recruitment and adopts an 'M2' phenotype associated with increased tissue fibrosis.
Keywords: Heart; Helminth infection; Macrophage.
Copyright © 2015 The Authors. Published by Elsevier GmbH.. All rights reserved.
Figures


References
-
- Epelman S., Lavine K.J., Beaudin A.E., Sojka D.K., Carrero J.A., Calderon B., Brija T., Gautier E.L., Ivanov S., Satpathy A.T., Schilling J.D., Schwendener R., Sergin I., Razani B., Forsberg E.C., Yokoyama W.M., Unanue E.R., Colonna M., Randolph G.J., Mann D.L. Embryonic and adult-derived resident cardiac macrophages are maintained through distinct mechanisms at steady state and during inflammation. Immunity. 2014;40:91–104. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
