

Development of twitching in sleeping infant mice depends on sensory experience
- PMID: 25702578
- PMCID: PMC4348337
- DOI: 10.1016/j.cub.2015.01.022
Development of twitching in sleeping infant mice depends on sensory experience
Erratum in
- Curr Biol. 2015 Jun 15;25(12):1672. Fritszch, Bernd [corrected to Fritzsch, Bernd]
Abstract
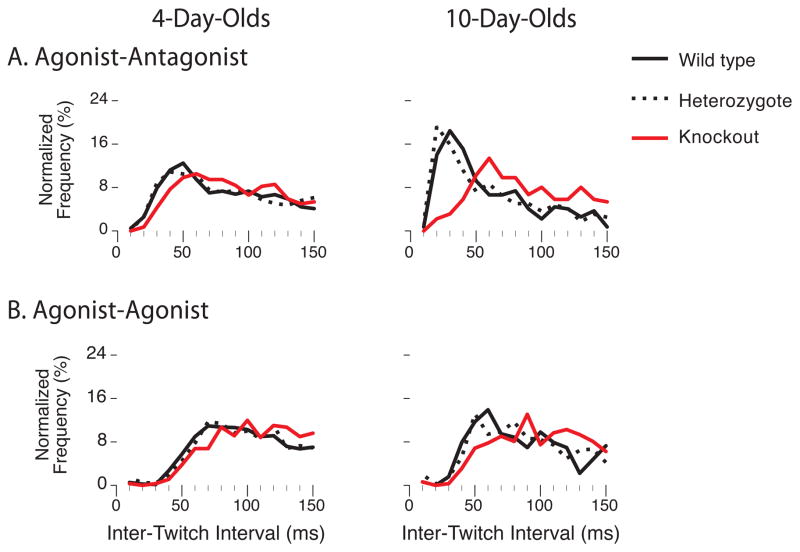
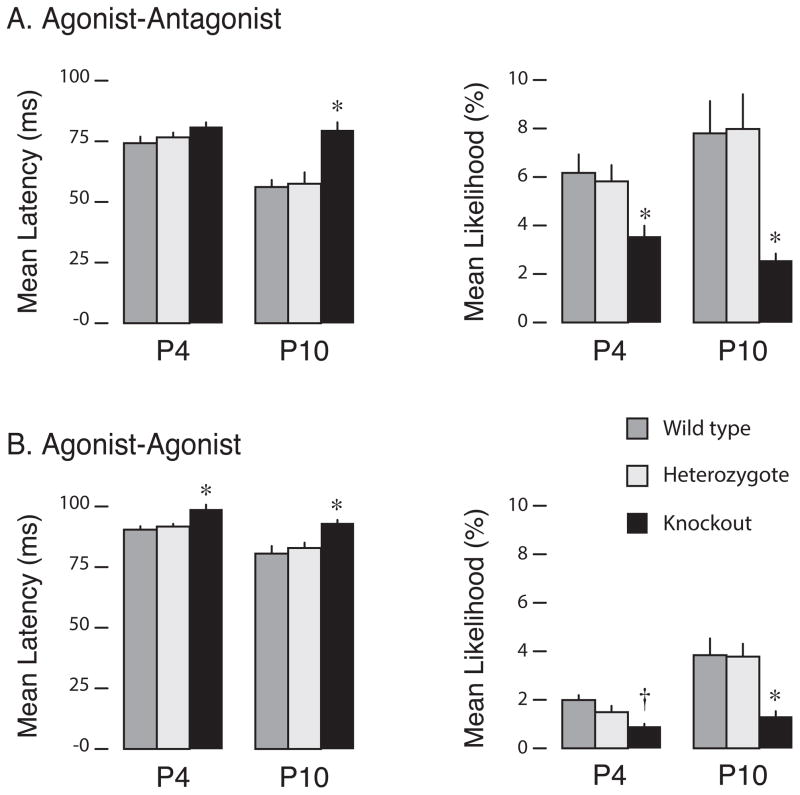
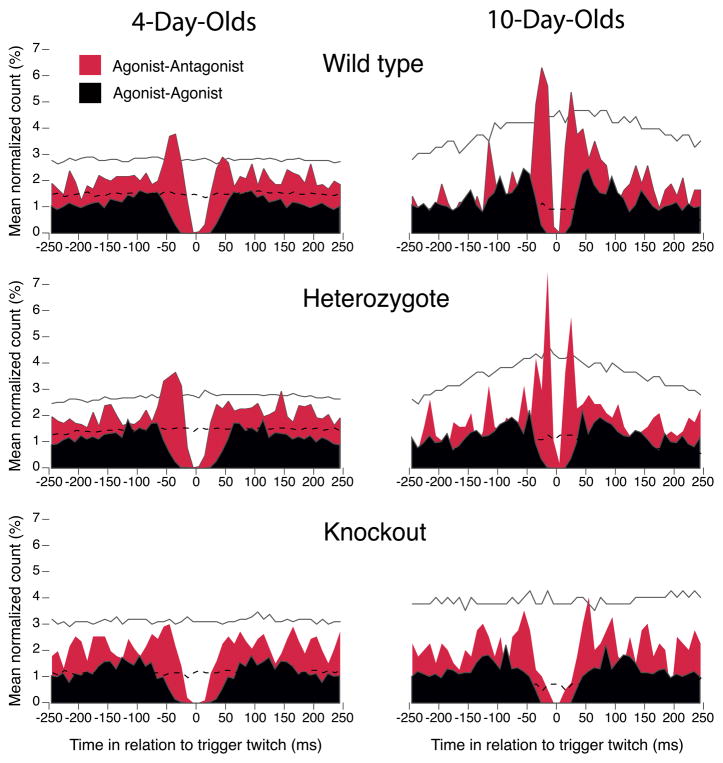
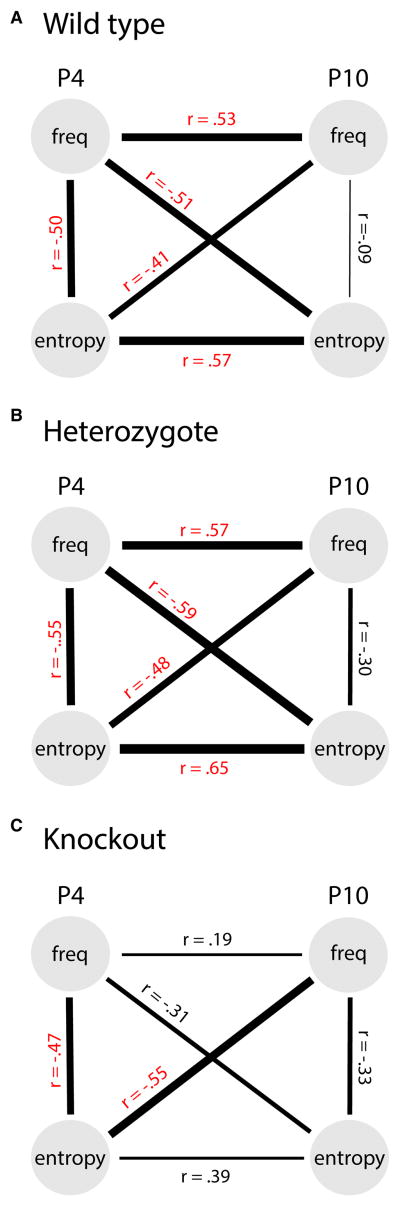
Myoclonic twitches are jerky movements that occur exclusively and abundantly during active (or REM) sleep in mammals, especially in early development [1-4]. In rat pups, limb twitches exhibit a complex spatiotemporal structure that changes across early development [5]. However, it is not known whether this developmental change is influenced by sensory experience, which is a prerequisite to the notion that sensory feedback from twitches not only activates sensorimotor circuits but modifies them [4]. Here, we investigated the contributions of proprioception to twitching in newborn ErbB2 conditional knockout mice that lack muscle spindles and grow up to exhibit dysfunctional proprioception [6-8]. High-speed videography of forelimb twitches unexpectedly revealed a category of reflex-like twitching-comprising an agonist twitch followed immediately by an antagonist twitch-that developed postnatally in wild-types/heterozygotes, but not in knockouts. Contrary to evidence from adults that spinal reflexes are inhibited during twitching [9-11], this finding suggests that twitches trigger the monosynaptic stretch reflex and, by doing so, contribute to its activity-dependent development [12-14]. Next, we assessed developmental changes in the frequency and organization (i.e., entropy) of more-complex, multi-joint patterns of twitching; again, wild-types/heterozygotes exhibited developmental changes in twitch patterning that were not seen in knockouts. Thus, targeted deletion of a peripheral sensor alters the normal development of local and global features of twitching, demonstrating that twitching is shaped by sensory experience. These results also highlight the potential use of twitching as a uniquely informative diagnostic tool for assessing the functional status of spinal and supraspinal circuits.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Spatiotemporal organization of myoclonic twitching in sleeping human infants.Dev Psychobiol. 2020 Sep;62(6):697-710. doi: 10.1002/dev.21954. Epub 2020 Feb 9. Dev Psychobiol. 2020. PMID: 32037557 Free PMC article.
-
Spatiotemporal structure of REM sleep twitching reveals developmental origins of motor synergies.Curr Biol. 2013 Nov 4;23(21):2100-9. doi: 10.1016/j.cub.2013.08.055. Epub 2013 Oct 17. Curr Biol. 2013. PMID: 24139739 Free PMC article.
-
Twitches emerge postnatally during quiet sleep in human infants and are synchronized with sleep spindles.Curr Biol. 2021 Aug 9;31(15):3426-3432.e4. doi: 10.1016/j.cub.2021.05.038. Epub 2021 Jun 17. Curr Biol. 2021. PMID: 34139191 Free PMC article.
-
Proprioception and myoclonus.Neurophysiol Clin. 2006 Sep-Dec;36(5-6):299-308. doi: 10.1016/j.neucli.2006.11.001. Epub 2006 Nov 30. Neurophysiol Clin. 2006. PMID: 17336774 Review.
-
Twitching in sensorimotor development from sleeping rats to robots.Curr Biol. 2013 Jun 17;23(12):R532-7. doi: 10.1016/j.cub.2013.04.075. Curr Biol. 2013. PMID: 23787051 Free PMC article. Review.
Cited by
-
A new view of "dream enactment" in REM sleep behavior disorder.Sleep Med Rev. 2016 Dec;30:34-42. doi: 10.1016/j.smrv.2015.12.002. Epub 2015 Dec 17. Sleep Med Rev. 2016. PMID: 26802823 Free PMC article. Review.
-
Phantom Limbs, Neuroprosthetics, and the Developmental Origins of Embodiment.Trends Neurosci. 2017 Oct;40(10):603-612. doi: 10.1016/j.tins.2017.07.003. Trends Neurosci. 2017. PMID: 28843655 Free PMC article. Review.
-
Twitches, Blinks, and Fidgets: Important Generators of Ongoing Neural Activity.Neuroscientist. 2019 Aug;25(4):298-313. doi: 10.1177/1073858418805427. Epub 2018 Oct 12. Neuroscientist. 2019. PMID: 30311838 Free PMC article. Review.
-
Spatiotemporal organization of myoclonic twitching in sleeping human infants.Dev Psychobiol. 2020 Sep;62(6):697-710. doi: 10.1002/dev.21954. Epub 2020 Feb 9. Dev Psychobiol. 2020. PMID: 32037557 Free PMC article.
-
Sensorimotor processing in the newborn rat red nucleus during active sleep.J Neurosci. 2015 May 27;35(21):8322-32. doi: 10.1523/JNEUROSCI.0564-15.2015. J Neurosci. 2015. PMID: 26019345 Free PMC article.
References
-
- Gramsbergen A, Schwartze P, Prechtl HFR. The postnatal development of behavioral states in the rat. Dev Psychobiol. 1970;3:267–280. - PubMed
-
- Jouvet-Mounier D, Astic L, Lacote D. Ontogenesis of the states of sleep in rat, cat, and guinea pig during the first postnatal month. Dev Psychobiol. 1970;2:216–239. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
