

Inverse size scaling of the nucleolus by a concentration-dependent phase transition
- PMID: 25702583
- PMCID: PMC4348177
- DOI: 10.1016/j.cub.2015.01.012
Inverse size scaling of the nucleolus by a concentration-dependent phase transition
Abstract
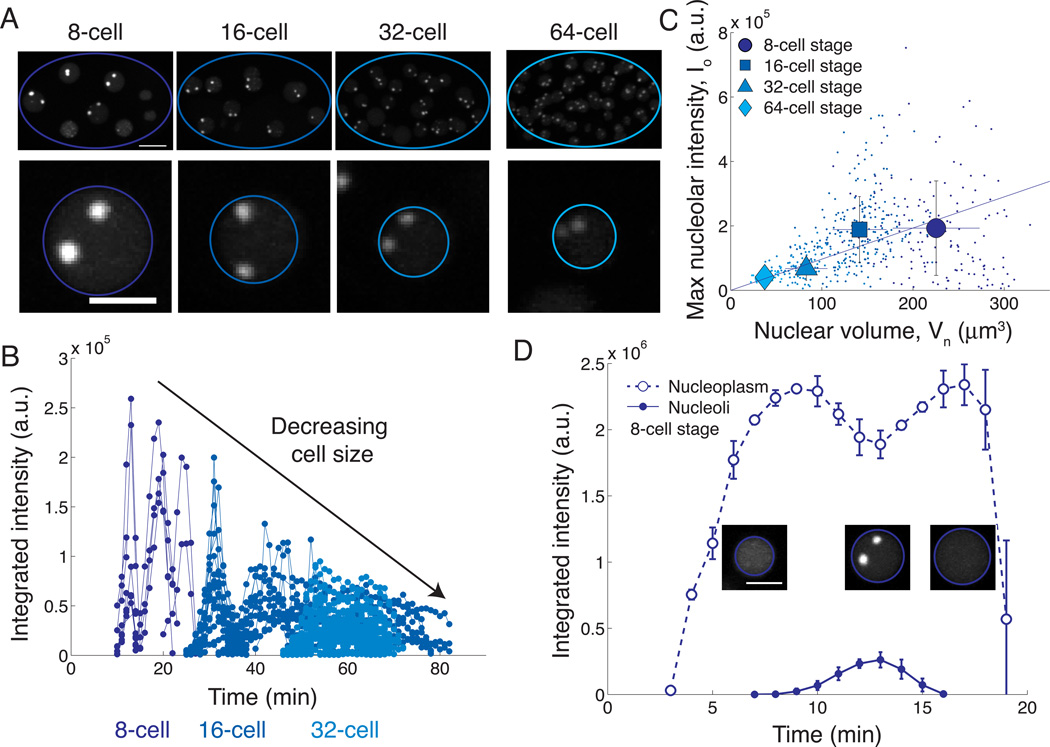
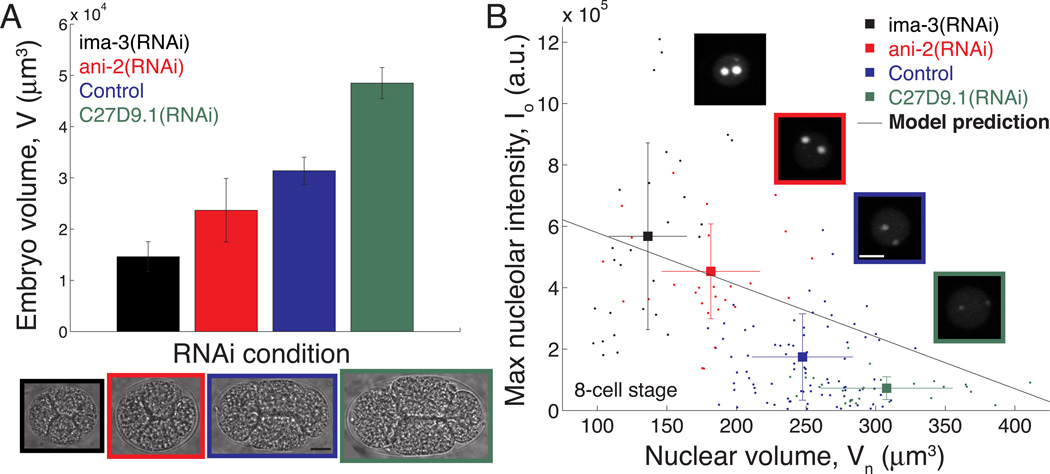
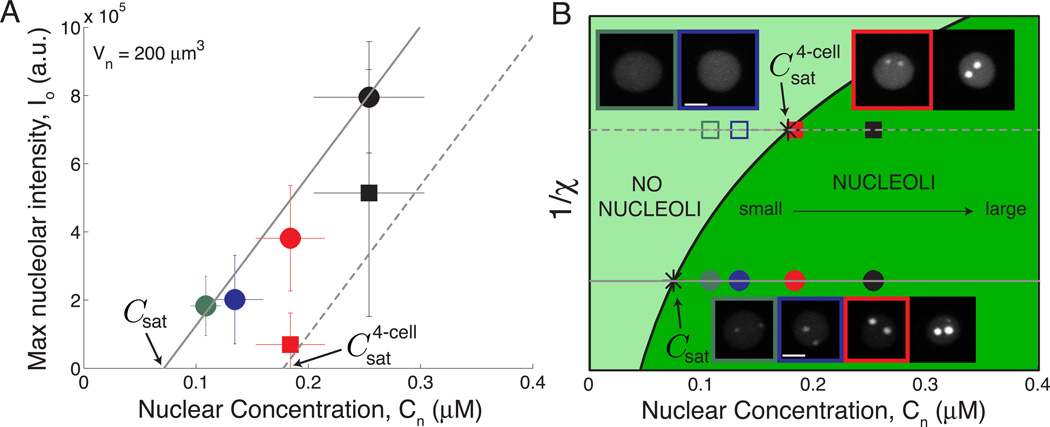
Just as organ size typically increases with body size, the size of intracellular structures changes as cells grow and divide. Indeed, many organelles, such as the nucleus [1, 2], mitochondria [3], mitotic spindle [4, 5], and centrosome [6], exhibit size scaling, a phenomenon in which organelle size depends linearly on cell size. However, the mechanisms of organelle size scaling remain unclear. Here, we show that the size of the nucleolus, a membraneless organelle important for cell-size homeostasis [7], is coupled to cell size by an intracellular phase transition. We find that nucleolar size directly scales with cell size in early C. elegans embryos. Surprisingly, however, when embryo size is altered, we observe inverse scaling: nucleolar size increases in small cells and decreases in large cells. We demonstrate that this seemingly contradictory result arises from maternal loading of a fixed number rather than a fixed concentration of nucleolar components, which condense into nucleoli only above a threshold concentration. Our results suggest that the physics of phase transitions can dictate whether an organelle assembles, and, if so, its size, providing a mechanistic link between organelle assembly and cell size. Since the nucleolus is known to play a key role in cell growth, this biophysical readout of cell size could provide a novel feedback mechanism for growth control.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures

Similar articles
-
Size scaling of nucleolus in Caenorhabditis elegans embryos.Biomed J. 2018 Oct;41(5):333-336. doi: 10.1016/j.bj.2018.07.003. Epub 2018 Nov 6. Biomed J. 2018. PMID: 30580798 Free PMC article.
-
RNA transcription modulates phase transition-driven nuclear body assembly.Proc Natl Acad Sci U S A. 2015 Sep 22;112(38):E5237-45. doi: 10.1073/pnas.1509317112. Epub 2015 Sep 8. Proc Natl Acad Sci U S A. 2015. PMID: 26351690 Free PMC article.
-
Centrosome size sets mitotic spindle length in Caenorhabditis elegans embryos.Curr Biol. 2010 Feb 23;20(4):353-8. doi: 10.1016/j.cub.2009.12.050. Epub 2010 Feb 4. Curr Biol. 2010. PMID: 20137951
-
Intracellular Scaling Mechanisms.Cold Spring Harb Perspect Biol. 2015 Aug 7;7(12):a019067. doi: 10.1101/cshperspect.a019067. Cold Spring Harb Perspect Biol. 2015. PMID: 26254310 Free PMC article. Review.
-
The nucleolus of Caenorhabditis elegans.J Biomed Biotechnol. 2012;2012:601274. doi: 10.1155/2012/601274. Epub 2012 Apr 19. J Biomed Biotechnol. 2012. PMID: 22577294 Free PMC article. Review.
Cited by
-
Size scaling of nucleolus in Caenorhabditis elegans embryos.Biomed J. 2018 Oct;41(5):333-336. doi: 10.1016/j.bj.2018.07.003. Epub 2018 Nov 6. Biomed J. 2018. PMID: 30580798 Free PMC article.
-
Component analysis of nucleolar protein compartments using Xenopus laevis oocytes.Dev Growth Differ. 2022 Aug;64(6):306-317. doi: 10.1111/dgd.12794. Epub 2022 Jun 12. Dev Growth Differ. 2022. PMID: 35607824 Free PMC article.
-
Recent advances in understanding nuclear size and shape.Nucleus. 2016 Apr 25;7(2):167-86. doi: 10.1080/19491034.2016.1162933. Epub 2016 Mar 10. Nucleus. 2016. PMID: 26963026 Free PMC article. Review.
-
At the Interface of Three Nucleic Acids: The Role of RNA-Binding Proteins and Poly(ADP-ribose) in DNA Repair.Acta Naturae. 2017 Apr-Jun;9(2):4-16. Acta Naturae. 2017. PMID: 28740723 Free PMC article.
-
Genetic mutations in RNA-binding proteins and their roles in ALS.Hum Genet. 2017 Sep;136(9):1193-1214. doi: 10.1007/s00439-017-1830-7. Epub 2017 Jul 31. Hum Genet. 2017. PMID: 28762175 Free PMC article. Review.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
