

Sensory detection of food rapidly modulates arcuate feeding circuits
- PMID: 25703096
- PMCID: PMC4373539
- DOI: 10.1016/j.cell.2015.01.033
Sensory detection of food rapidly modulates arcuate feeding circuits
Abstract
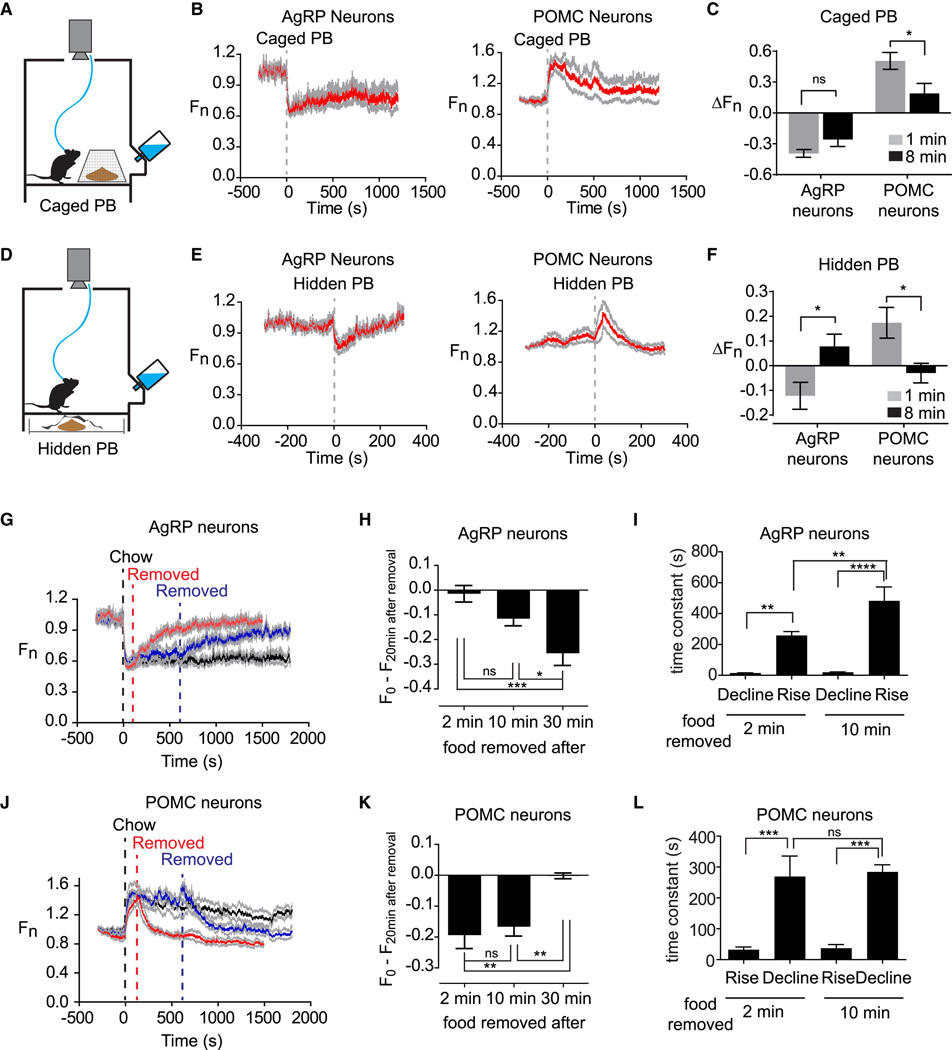
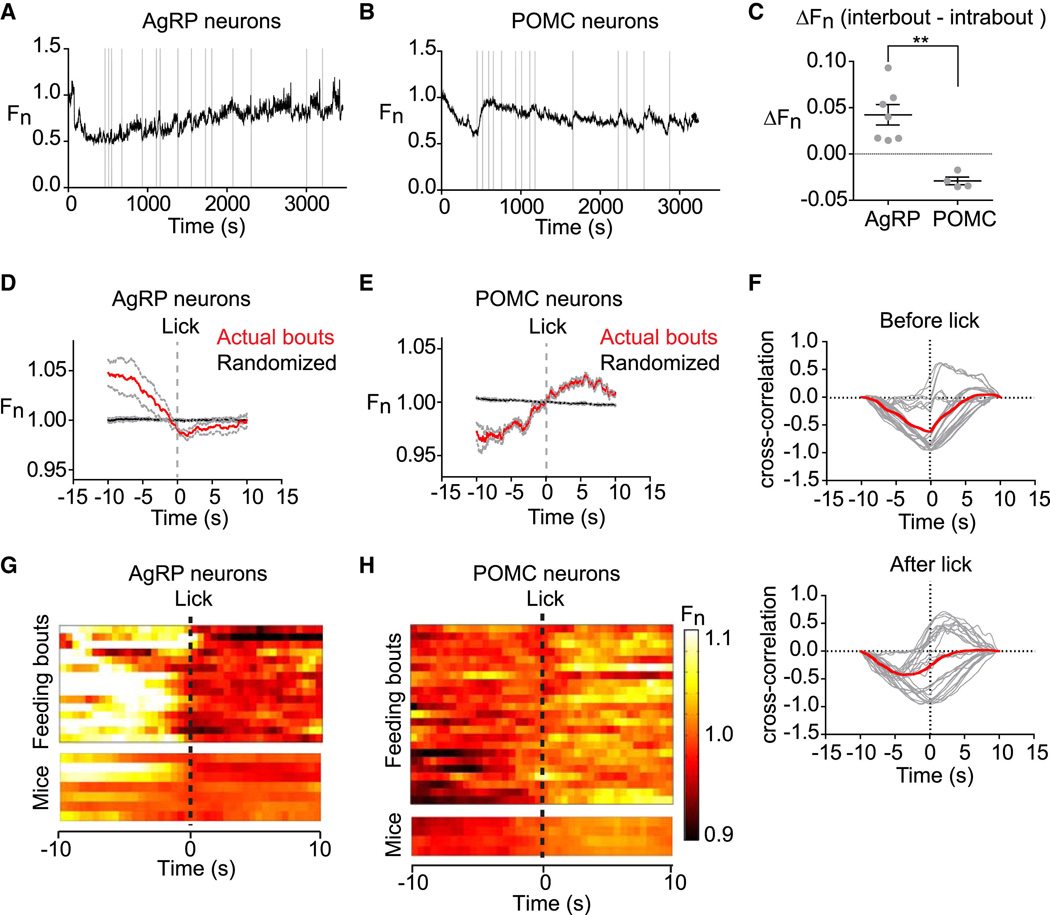
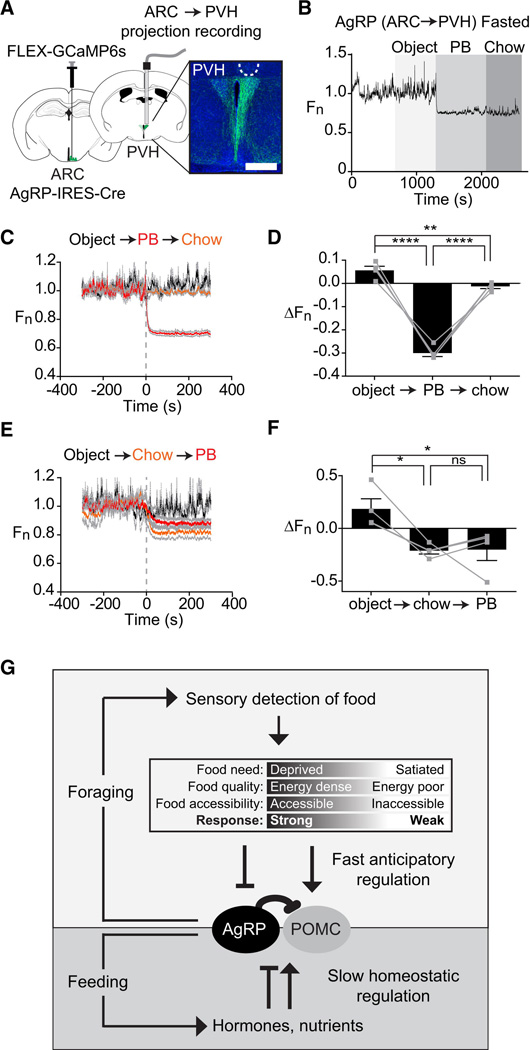
Hunger is controlled by specialized neural circuits that translate homeostatic needs into motivated behaviors. These circuits are under chronic control by circulating signals of nutritional state, but their rapid dynamics on the timescale of behavior remain unknown. Here, we report optical recording of the natural activity of two key cell types that control food intake, AgRP and POMC neurons, in awake behaving mice. We find unexpectedly that the sensory detection of food is sufficient to rapidly reverse the activation state of these neurons induced by energy deficit. This rapid regulation is cell-type specific, modulated by food palatability and nutritional state, and occurs before any food is consumed. These data reveal that AgRP and POMC neurons receive real-time information about the availability of food in the external world, suggesting a primary role for these neurons in controlling appetitive behaviors such as foraging that promote the discovery of food.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
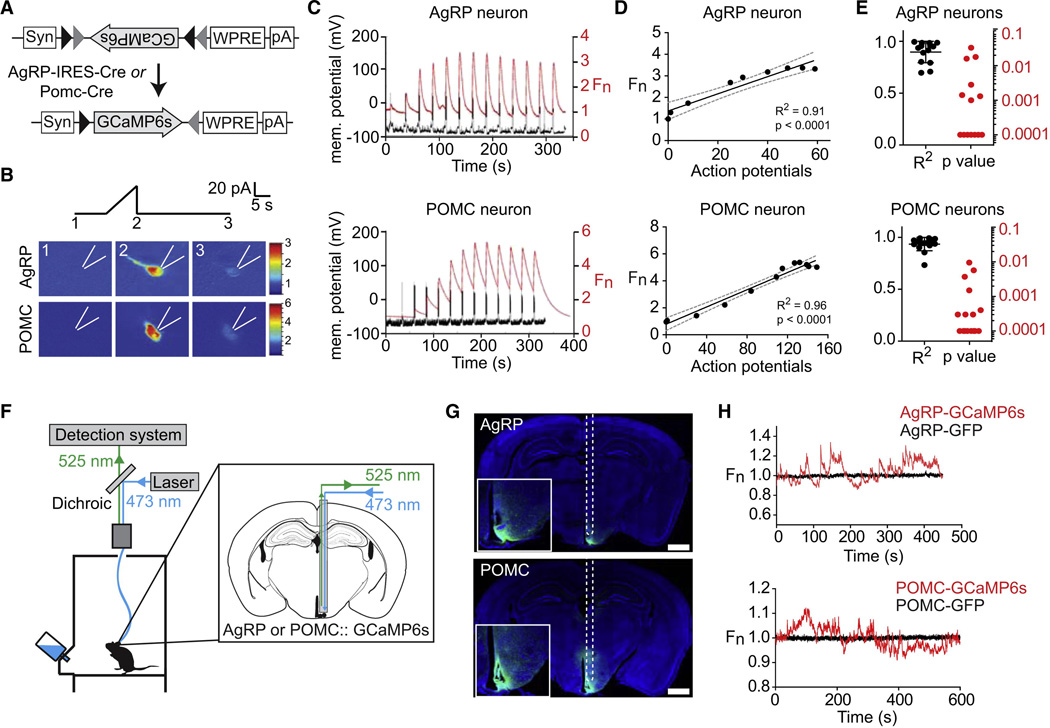
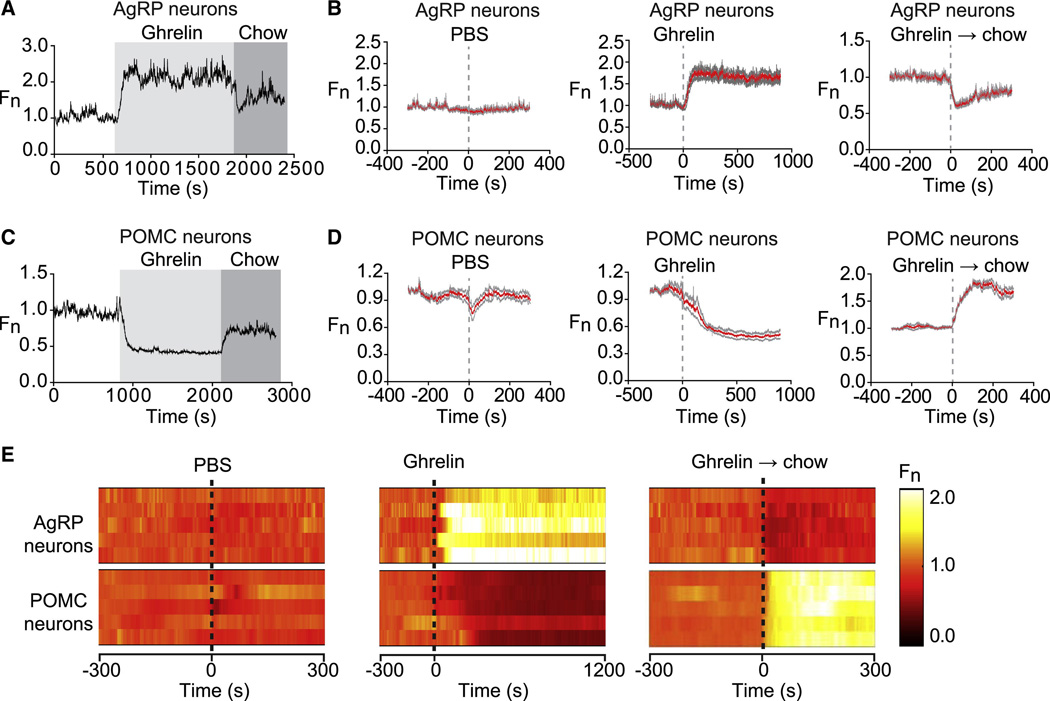
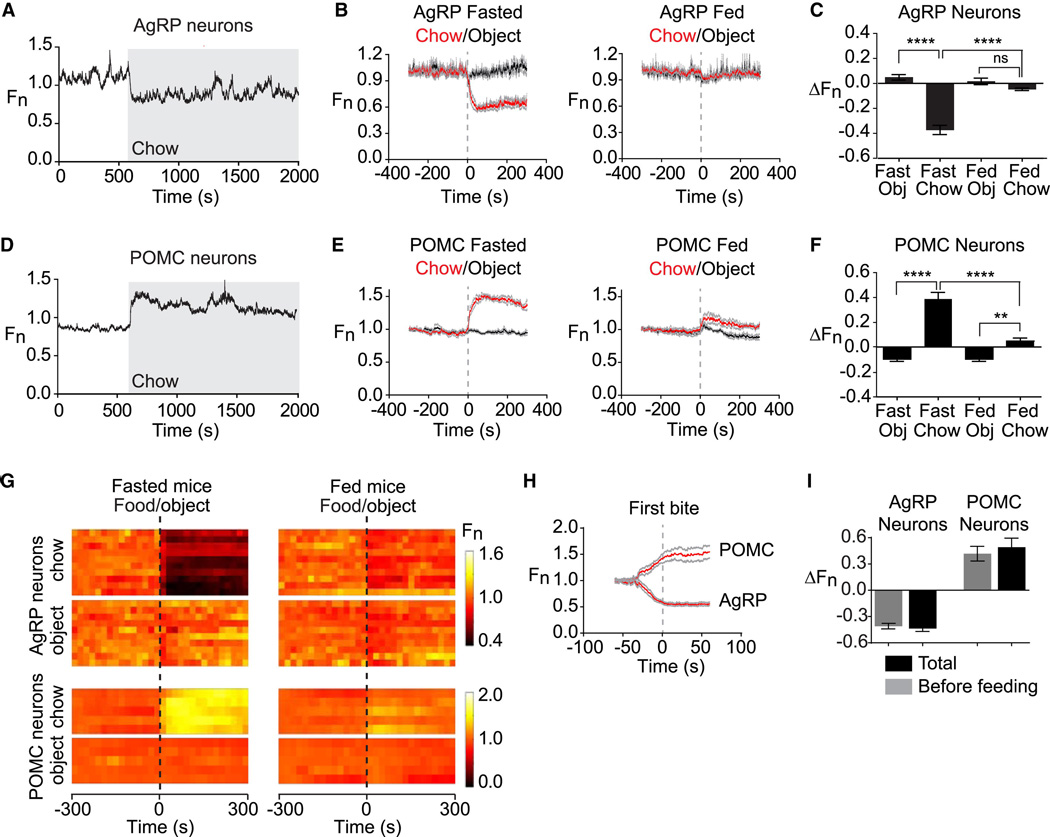
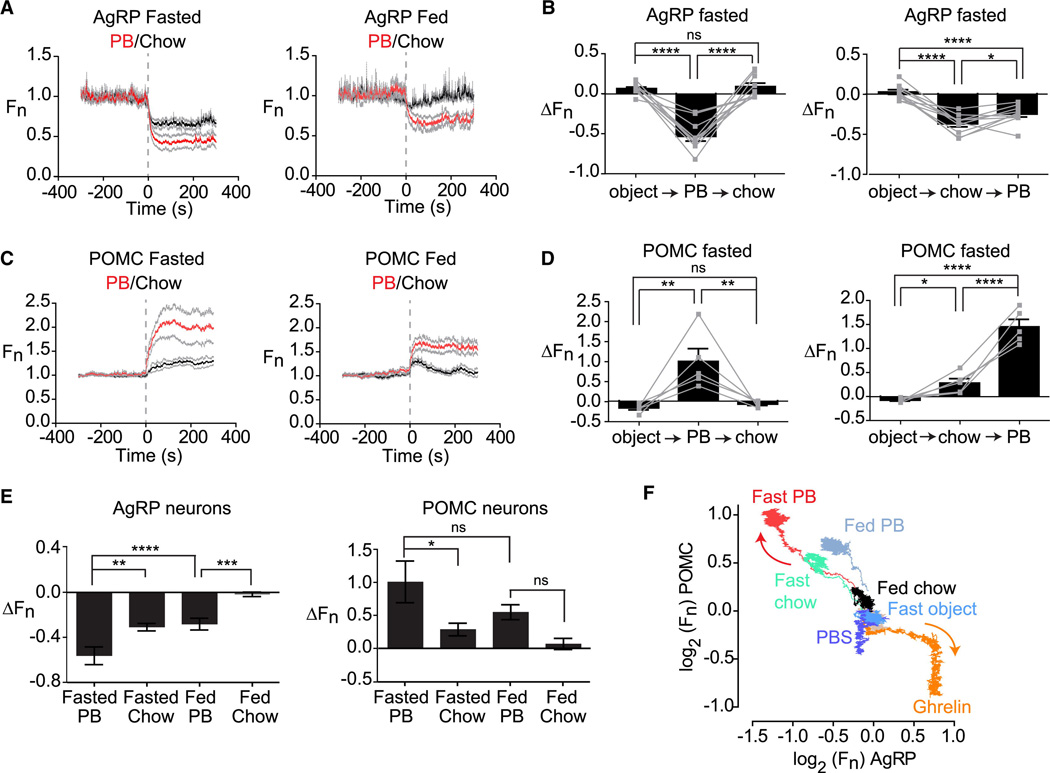
Comment in
-
The hunger games.Cell. 2015 Feb 26;160(5):805-806. doi: 10.1016/j.cell.2015.02.028. Cell. 2015. PMID: 25723156
References
-
- Blouet C, Schwartz GJ. Hypothalamic nutrient sensing in the control of energy homeostasis. Behavioural brain research. 2010;209:1–12. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
