

A conserved activation cluster is required for allosteric communication in HtrA-family proteases
- PMID: 25703375
- PMCID: PMC4351158
- DOI: 10.1016/j.str.2015.01.012
A conserved activation cluster is required for allosteric communication in HtrA-family proteases
Abstract
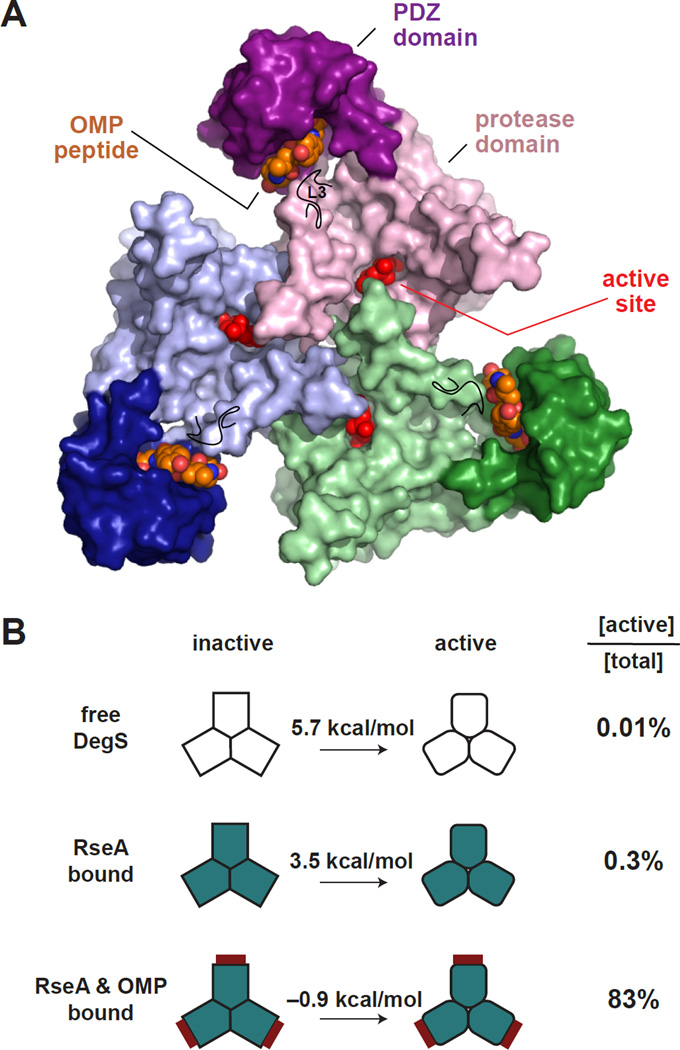
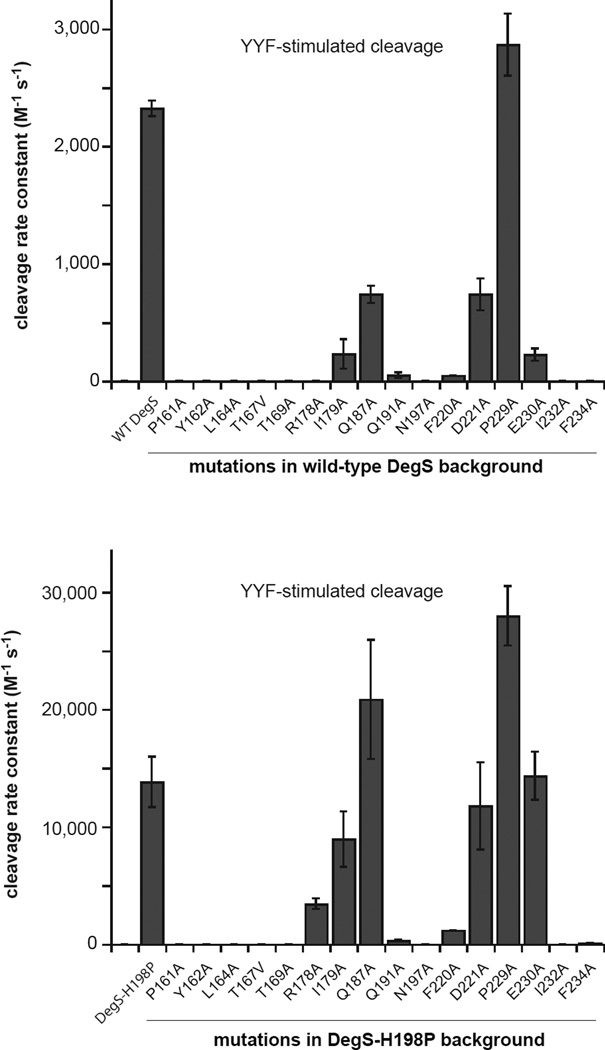
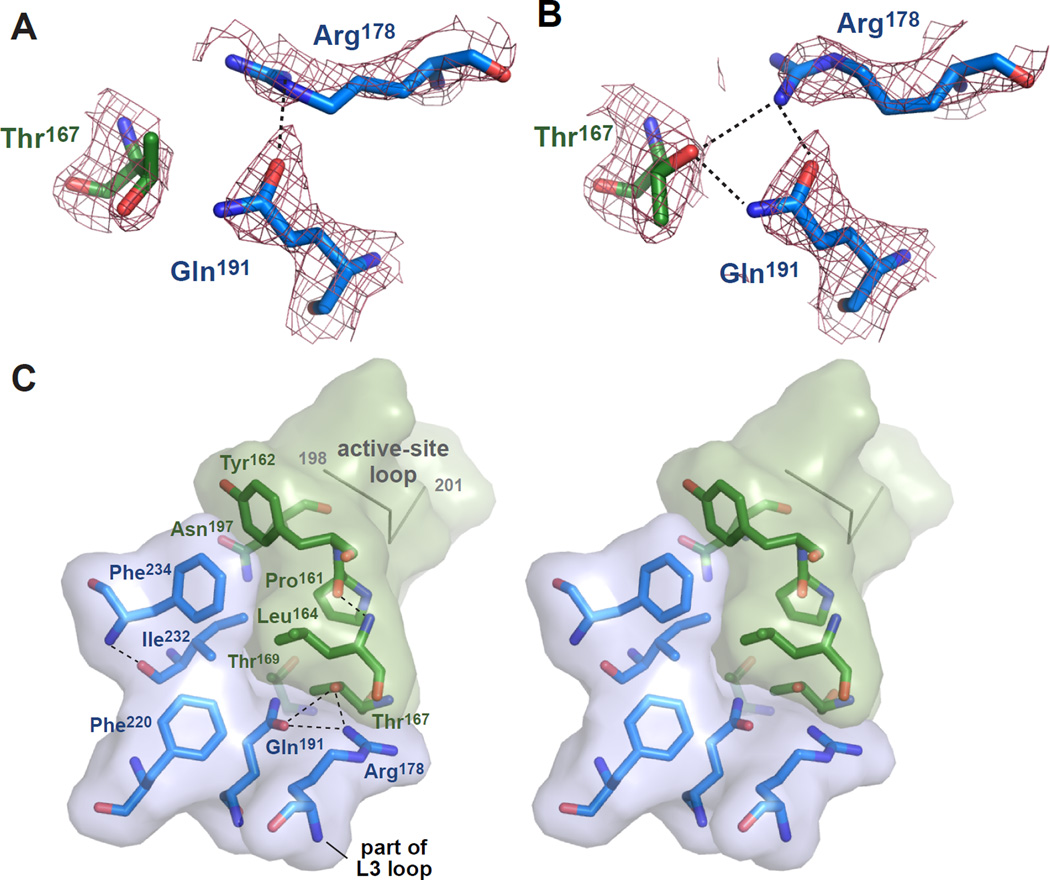
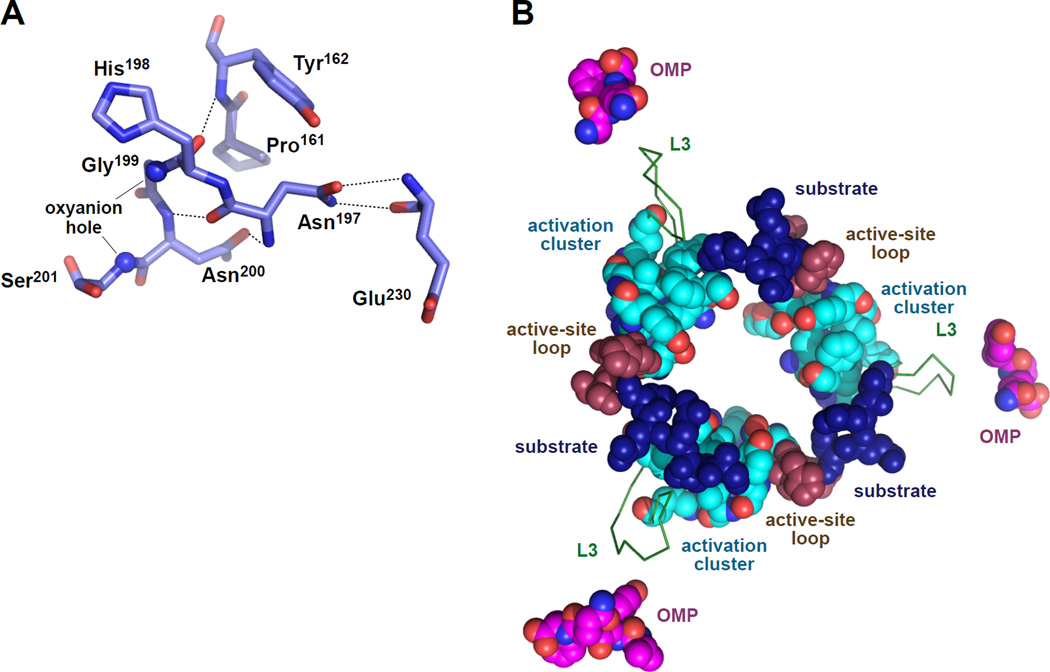
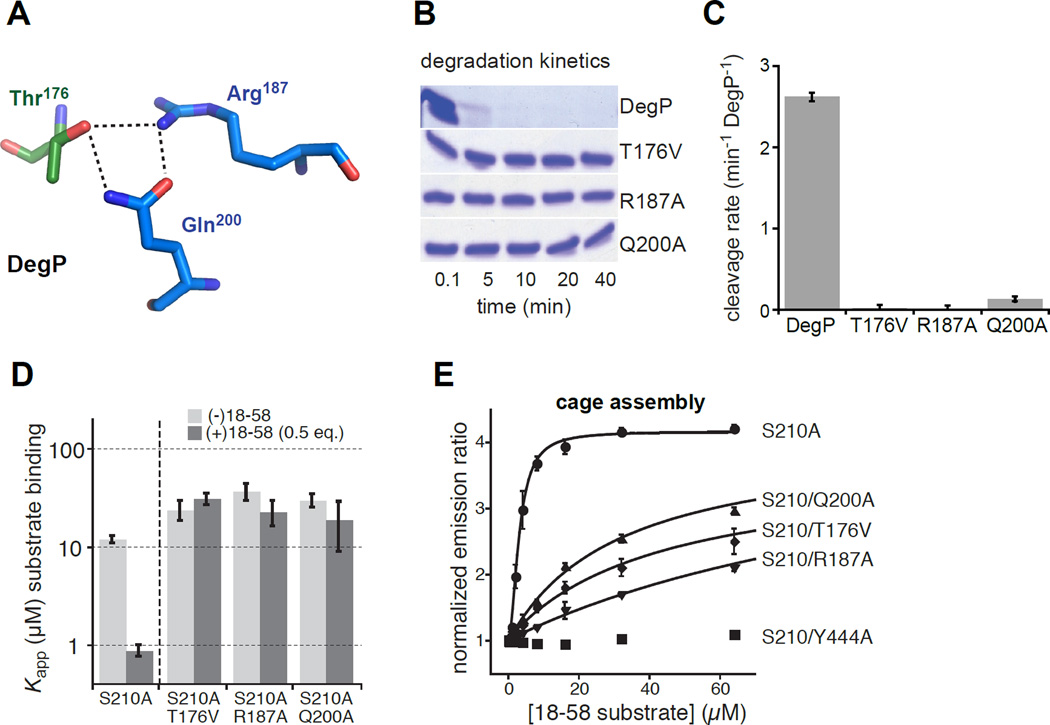
In E. coli, outer-membrane stress causes a transcriptional response through a signaling cascade initiated by DegS cleavage of a transmembrane antisigma factor. Each subunit of DegS, an HtrA-family protease, contains a protease domain and a PDZ domain. The trimeric protease domain is autoinhibited by the unliganded PDZ domains. Allosteric activation requires binding of unassembled outer-membrane proteins (OMPs) to the PDZ domains and protein substrate binding. Here, we identify a set of DegS residues that cluster together at subunit-subunit interfaces in the trimer, link the active sites and substrate binding sites, and are crucial for stabilizing the active enzyme conformation in response to OMP signaling. These residues are conserved across the HtrA-protease family, including orthologs linked to human disease, supporting a common mechanism of allosteric activation. Indeed, mutation of residues at homologous positions in the DegP quality-control protease also eliminates allosteric activation.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures

References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
