

Interferons and viruses: an evolutionary arms race of molecular interactions
- PMID: 25704559
- PMCID: PMC4384471
- DOI: 10.1016/j.it.2015.01.004
Interferons and viruses: an evolutionary arms race of molecular interactions
Abstract
Over half a century has passed since interferons (IFNs) were discovered and shown to inhibit virus infection in cultured cells. Since then, researchers have steadily brought to light the molecular details of IFN signaling, catalogued their pleiotropic effects on cells, and harnessed their therapeutic potential for a variety of maladies. While advances have been plentiful, several fundamental questions have yet to be answered and much complexity remains to be unraveled. We explore the current knowledge surrounding four main questions: are type I IFN subtypes differentially produced in response to distinct pathogens? How are IFN subtypes distinguished by cells? What are the mechanisms and consequences of viral antagonism? Lastly, how can the IFN response be harnessed to improve vaccine efficacy?
Keywords: innate immunity; interferon subtype; interferons; vaccine adjuvants.; viral antagonism; viruses.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures


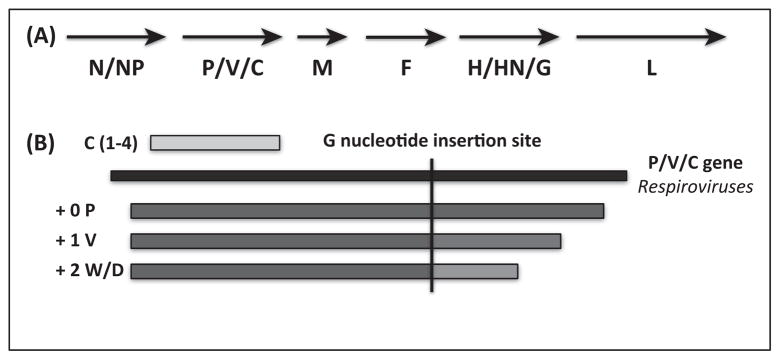
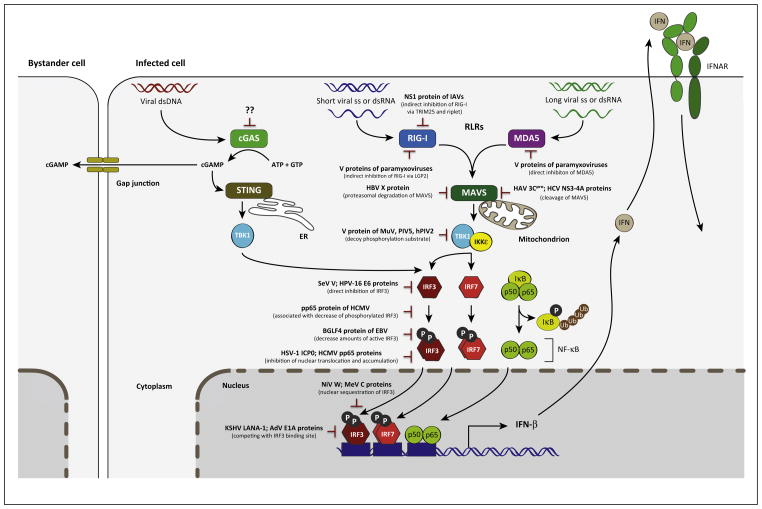

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
