

A distinct mechanism of vascular lumen formation in Xenopus requires EGFL7
- PMID: 25705891
- PMCID: PMC4338030
- DOI: 10.1371/journal.pone.0116086
A distinct mechanism of vascular lumen formation in Xenopus requires EGFL7
Abstract
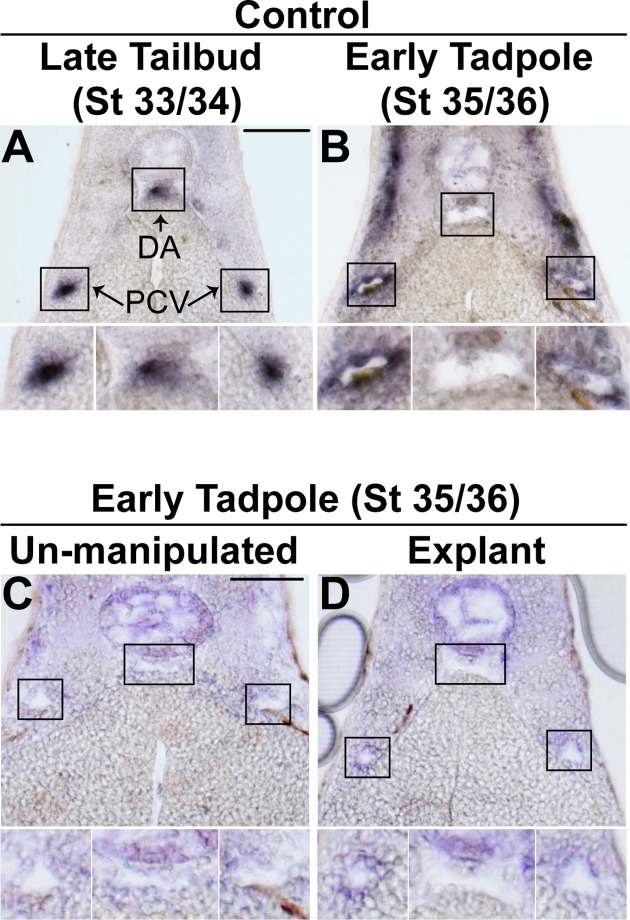
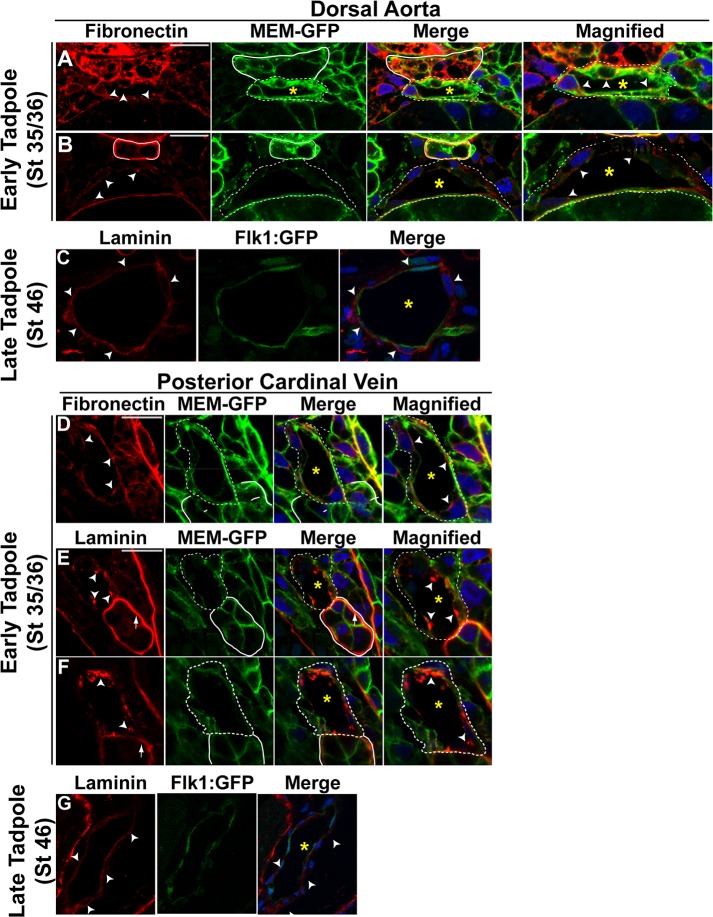
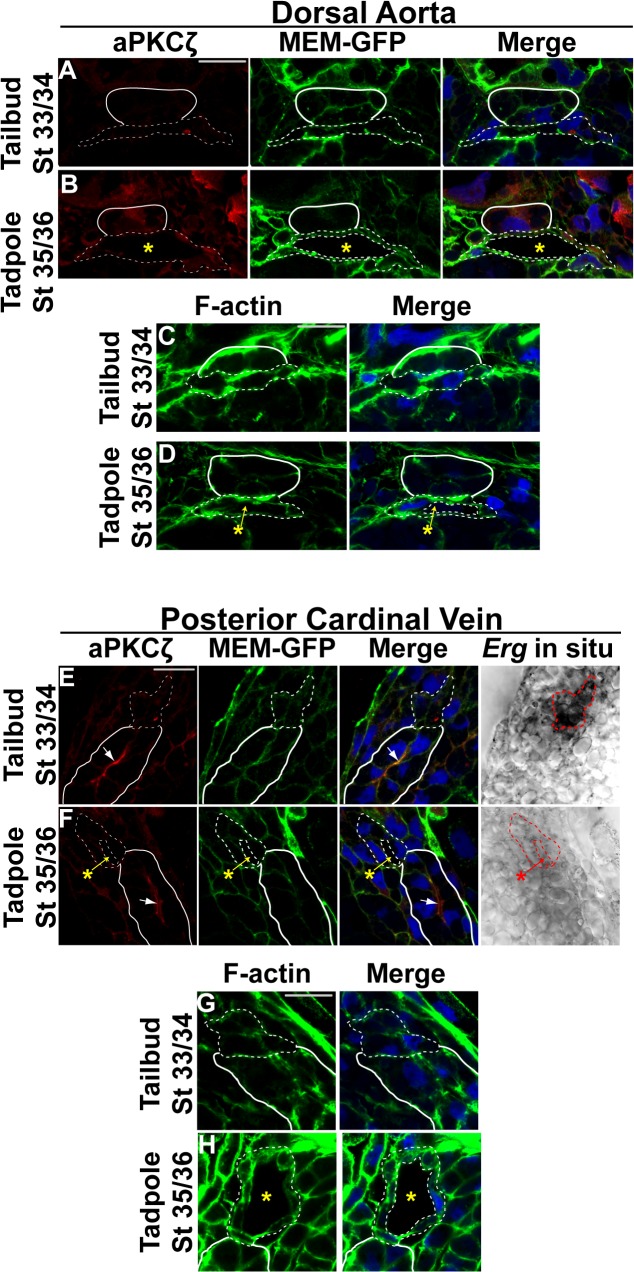
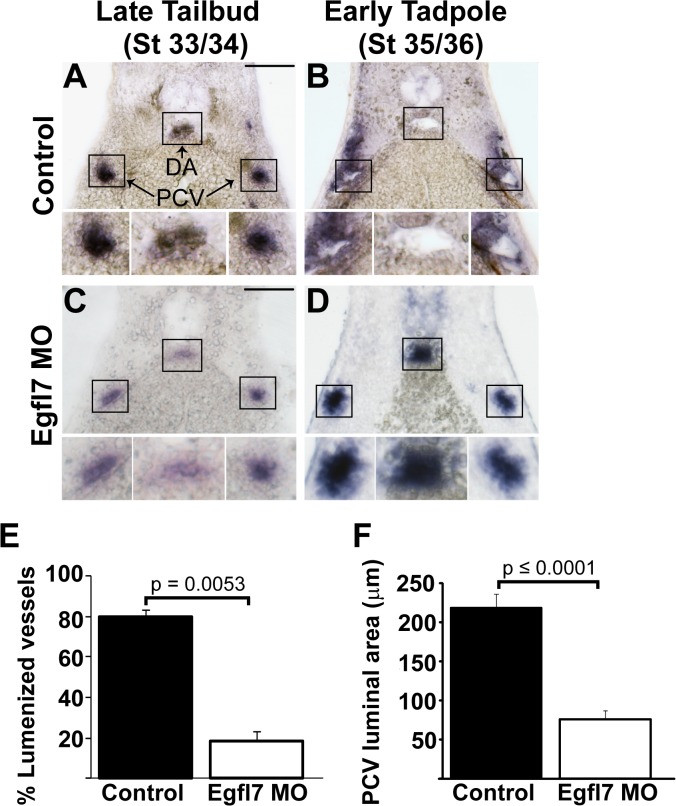
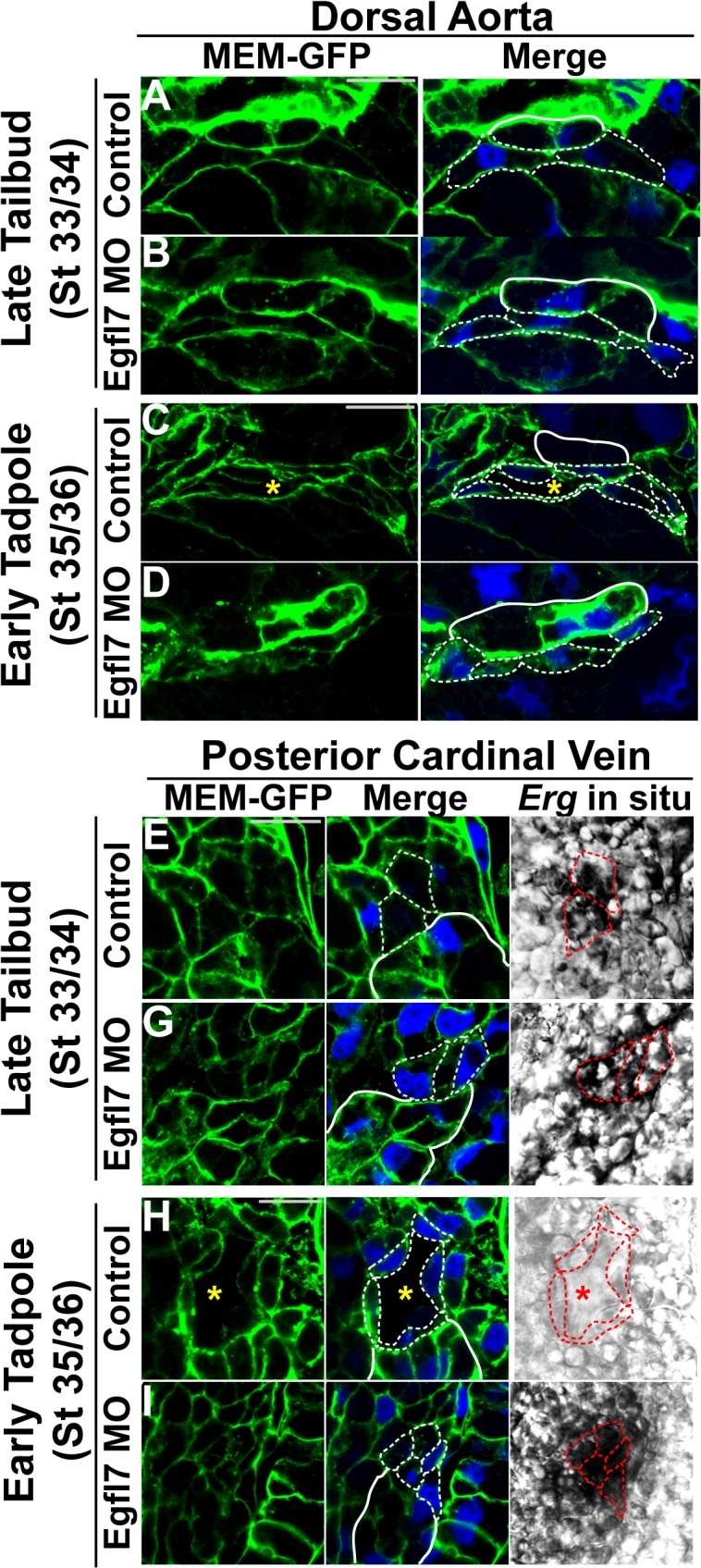
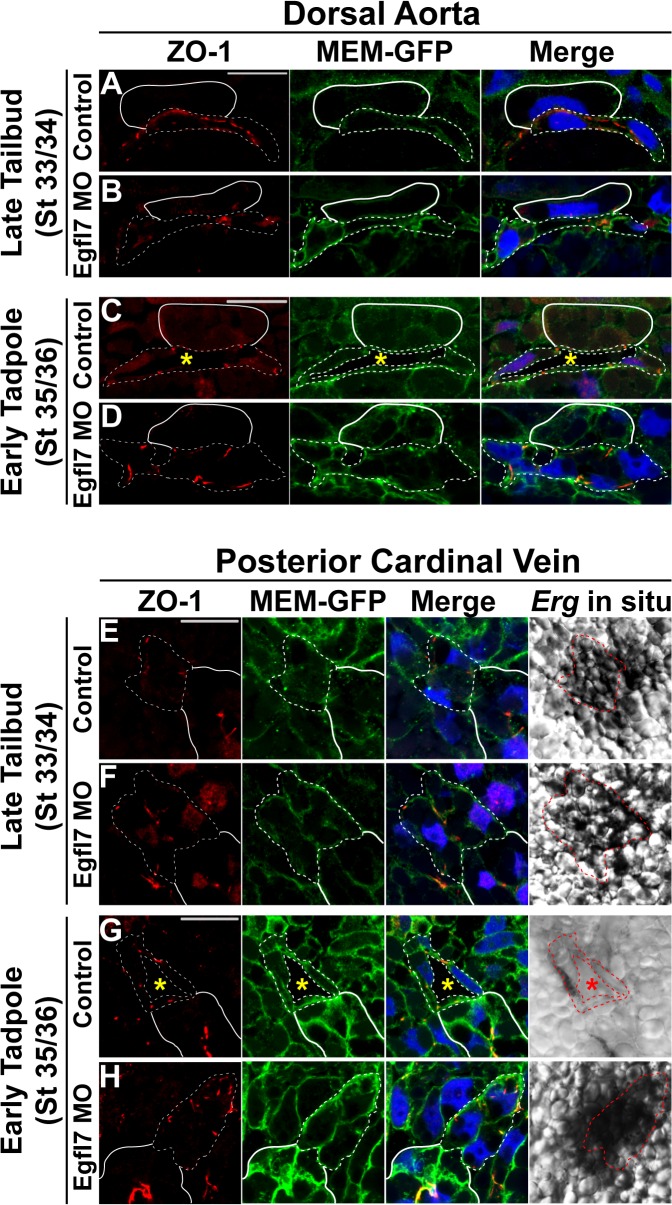
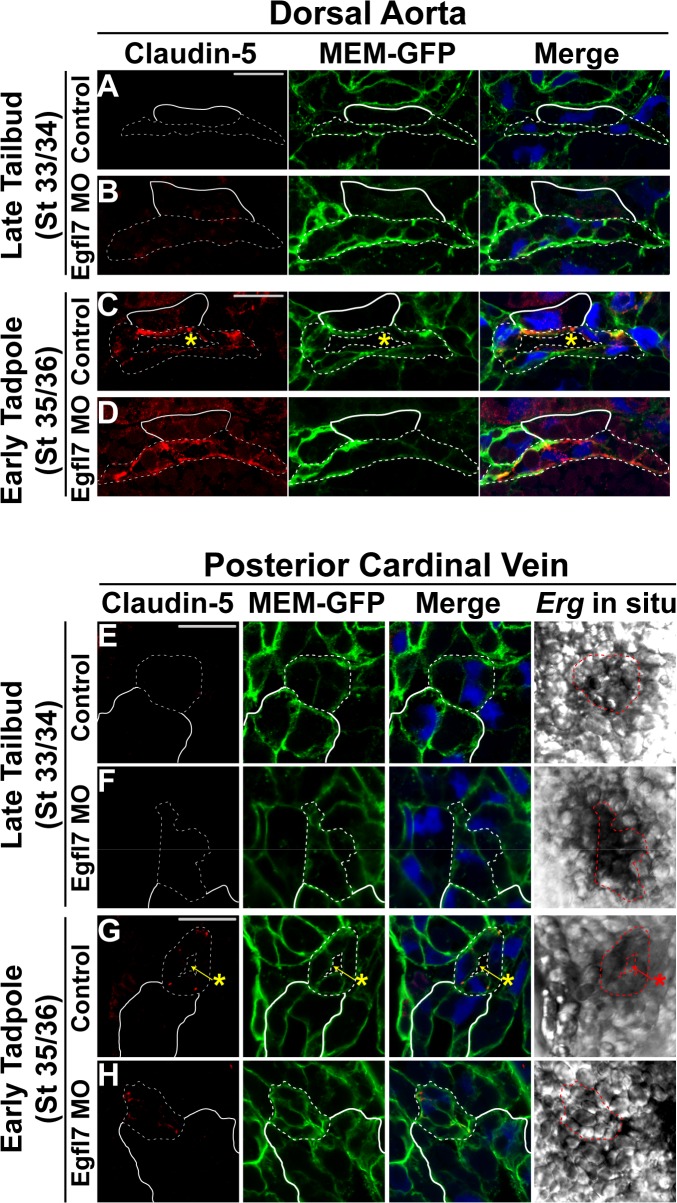
During vertebrate blood vessel development, lumen formation is the critical process by which cords of endothelial cells transition into functional tubular vessels. Here, we use Xenopus embryos to explore the cellular and molecular mechanisms underlying lumen formation of the dorsal aorta and the posterior cardinal veins, the primary major vessels that arise via vasculogenesis within the first 48 hours of life. We demonstrate that endothelial cells are initially found in close association with one another through the formation of tight junctions expressing ZO-1. The emergence of vascular lumens is characterized by elongation of endothelial cell shape, reorganization of junctions away from the cord center to the periphery of the vessel, and onset of Claudin-5 expression within tight junctions. Furthermore, unlike most vertebrate vessels that exhibit specialized apical and basal domains, we show that early Xenopus vessels are not polarized. Moreover, we demonstrate that in embryos depleted of the extracellular matrix factor Epidermal Growth Factor-Like Domain 7 (EGFL7), an evolutionarily conserved factor associated with vertebrate vessel development, vascular lumens fail to form. While Claudin-5 localizes to endothelial tight junctions of EGFL7-depleted embryos in a timely manner, endothelial cells of the aorta and veins fail to undergo appropriate cell shape changes or clear junctions from the cell-cell contact. Taken together, we demonstrate for the first time the mechanisms by which lumens are generated within the major vessels in Xenopus and implicate EGFL7 in modulating cell shape and cell-cell junctions to drive proper lumen morphogenesis.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
