

Robust and fragile aspects of cortical blood flow in relation to the underlying angioarchitecture
- PMID: 25705966
- PMCID: PMC4393797
- DOI: 10.1111/micc.12195
Robust and fragile aspects of cortical blood flow in relation to the underlying angioarchitecture
Abstract
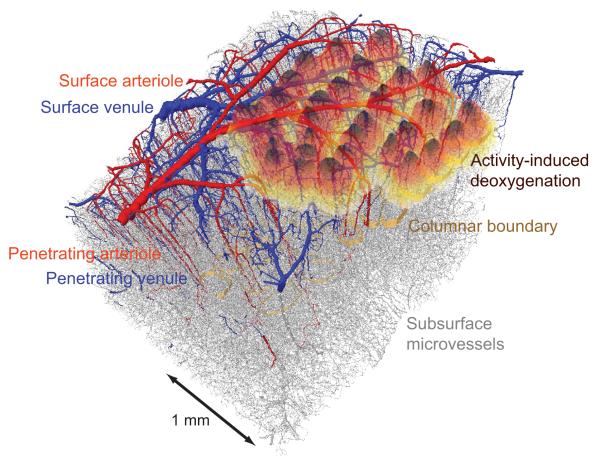
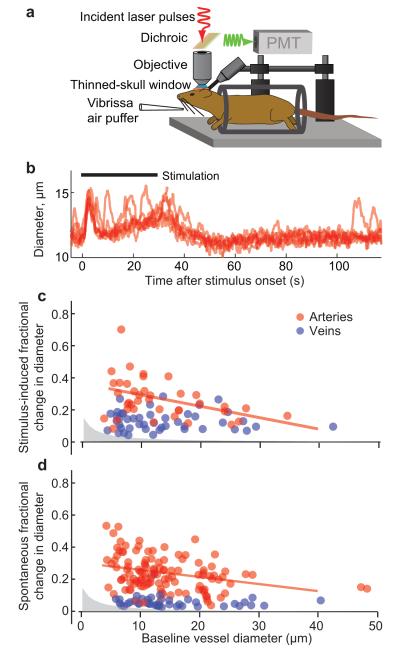
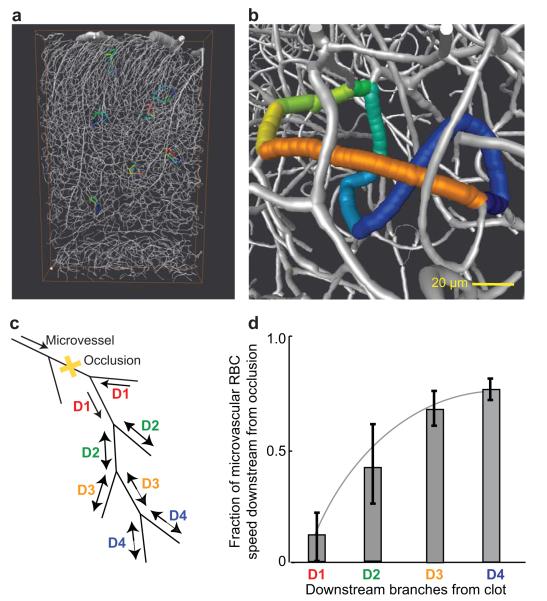
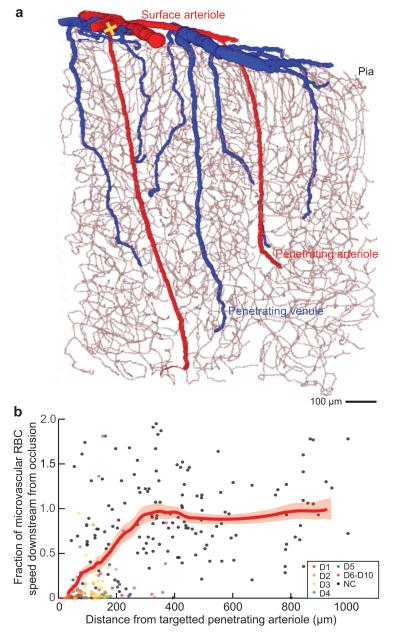
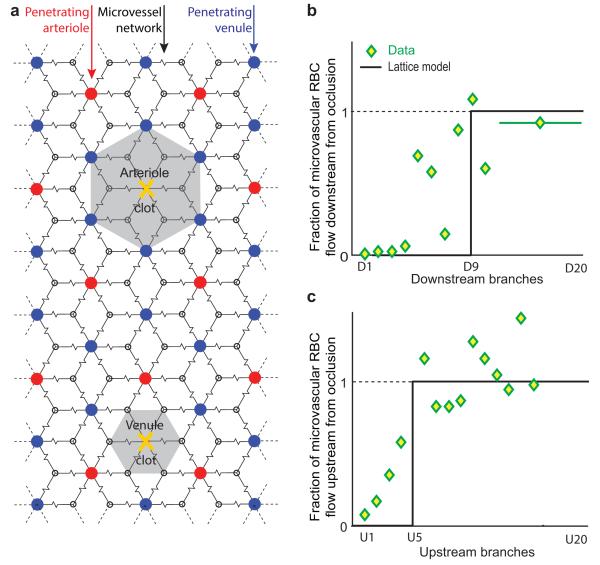
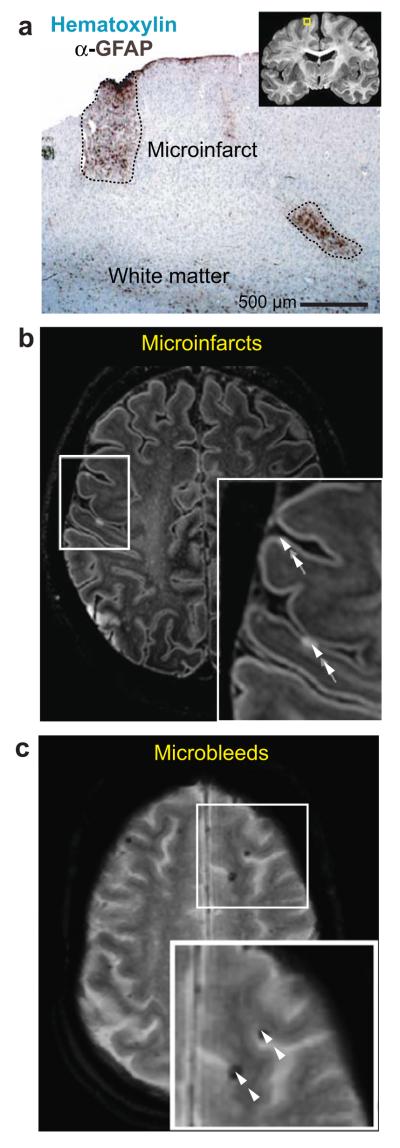
We review the organizational principles of the cortical vasculature and the underlying patterns of blood flow under normal conditions and in response to occlusion of single vessels. The cortex is sourced by a two-dimensional network of pial arterioles that feeds a three-dimensional network of subsurface microvessels in close proximity to neurons and glia. Blood flow within the surface and subsurface networks is largely insensitive to occlusion of a single vessel within either network. However, the penetrating arterioles that connect the pial network to the subsurface network are bottlenecks to flow; occlusion of even a single penetrating arteriole results in the death of a 500 μm diameter cylinder of cortical tissue despite the potential for collateral flow through microvessels. This pattern of flow is consistent with that calculated from a full reconstruction of the angioarchitecture. Conceptually, collateral flow is insufficient to compensate for the occlusion of a penetrating arteriole because penetrating venules act as shunts of blood that flows through collaterals. Future directions that stem from the analysis of the angioarchitecture concern cellular-level issues, in particular the regulation of blood flow within the subsurface microvascular network, and system-level issues, in particular the role of penetrating arteriole occlusions in human cognitive impairment.
Keywords: imaging; microvessels; penetrating vessels; pial vessels; rodent.
© 2015 John Wiley & Sons Ltd.
Figures




References
-
- Kleinfeld D, Denk W. Two-photon imaging of neocortical microcirculation. In: Yuste R, Lanni F, Konnerth A, editors. Imaging Neurons: A Laboratory Manual. Cold Spring Harbor Laboratory Press; Cold Spring Harbor: 1999. pp. 23.1–23.15.
-
- Tsai PS, Kleinfeld D. In vivo two-photon laser scanning microscopy with concurrent plasma-mediated ablation: Principles and hardware realization. In: Frostig RD, editor. Methods for In Vivo Optical Imaging. 2nd edition CRC Press; Boca Raton: 2009. pp. 59–115. - PubMed
-
- Kleinfeld D, Mitra PP. Applications of spectral methods in functional brain imaging. In: Yuste R, editor. Imaging: A Laboratory Manual. Cold Spring Harbor Laboratory Press; New York: 2011. pp. 12.1–12.7.
-
- Shih AY, Mateo C, Drew PJ, Tsai PS, Kleinfeld D. A polished and reinforced thinned skull window for long-term imaging and optical manipulation of the mouse cortex. Journal of Visualized Experiments. 2012 http://www.jove.com/video/3742 - PMC - PubMed
Publication types
MeSH terms
Grants and funding
- P20GM12345/GM/NIGMS NIH HHS/United States
- NS059832/NS/NINDS NIH HHS/United States
- DP1 OD006831/OD/NIH HHS/United States
- R21 MH072570/MH/NIMH NIH HHS/United States
- EB003832/EB/NIBIB NIH HHS/United States
- R21 NS059832/NS/NINDS NIH HHS/United States
- UL1TR000062/TR/NCATS NIH HHS/United States
- MH72570/MH/NIMH NIH HHS/United States
- RR021907/RR/NCRR NIH HHS/United States
- NS085402/NS/NINDS NIH HHS/United States
- R01 EB000790/EB/NIBIB NIH HHS/United States
- R01 NS057198/NS/NINDS NIH HHS/United States
- OD006831/OD/NIH HHS/United States
- R01 DA029706/DA/NIDA NIH HHS/United States
- UL1 TR000062/TR/NCATS NIH HHS/United States
- R01 EB003832/EB/NIBIB NIH HHS/United States
- NS082097/NS/NINDS NIH HHS/United States
- DP1 NS082097/NS/NINDS NIH HHS/United States
- R01 MH085499/MH/NIMH NIH HHS/United States
- R01 NS041096/NS/NINDS NIH HHS/United States
- R21 NS085402/NS/NINDS NIH HHS/United States
- R21 RR021907/RR/NCRR NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
