

SMAD signaling and redox imbalance cooperate to induce prostate cancer cell dormancy
- PMID: 25706341
- PMCID: PMC4615032
- DOI: 10.1080/15384101.2015.1014145
SMAD signaling and redox imbalance cooperate to induce prostate cancer cell dormancy
Abstract
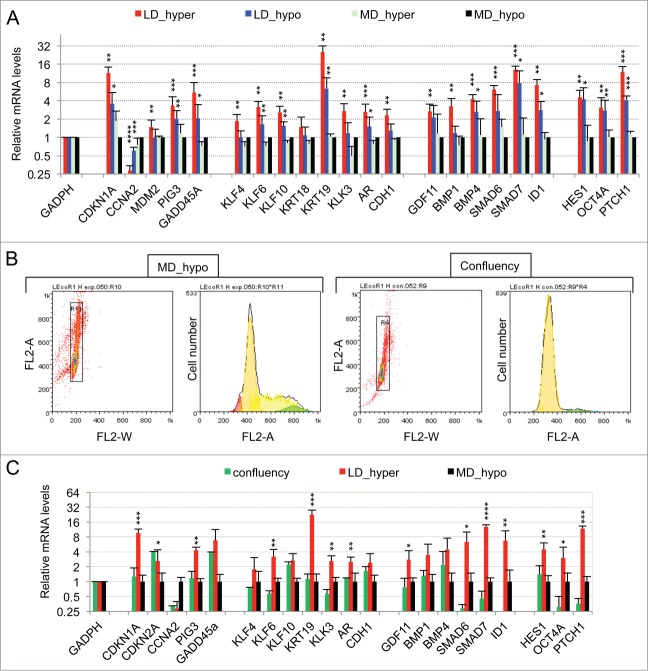
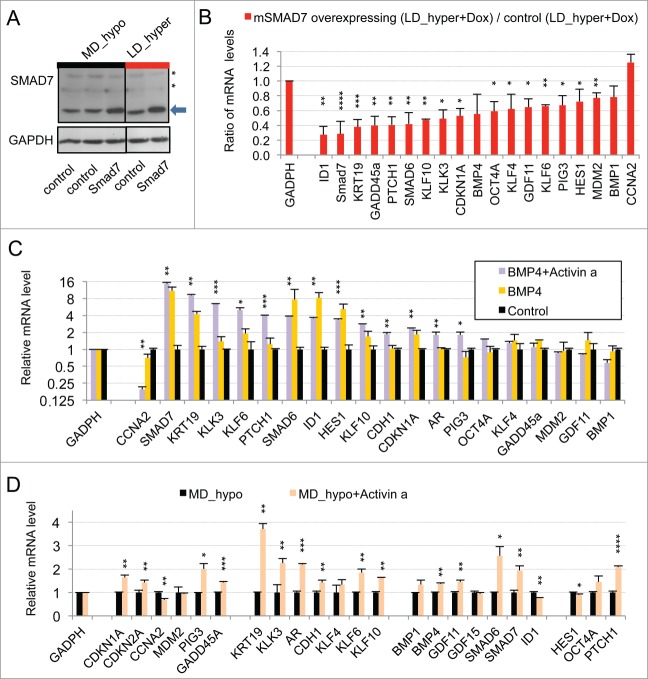
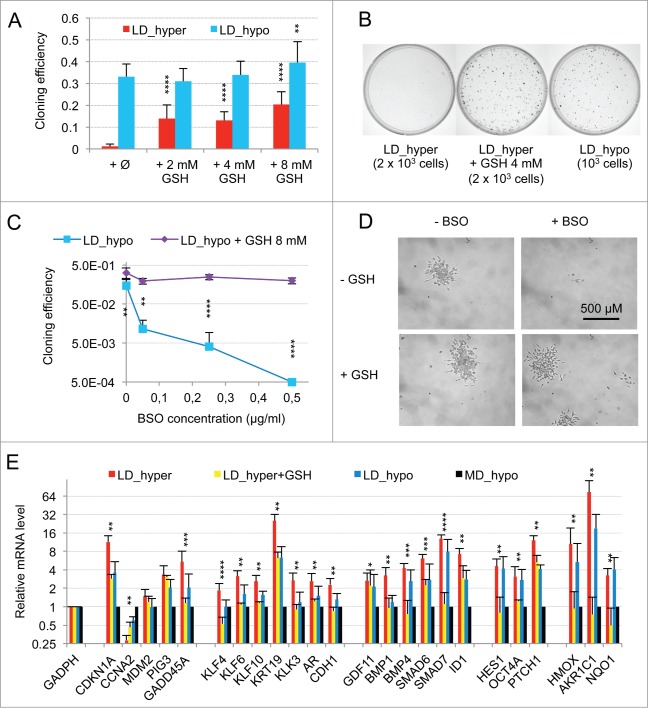
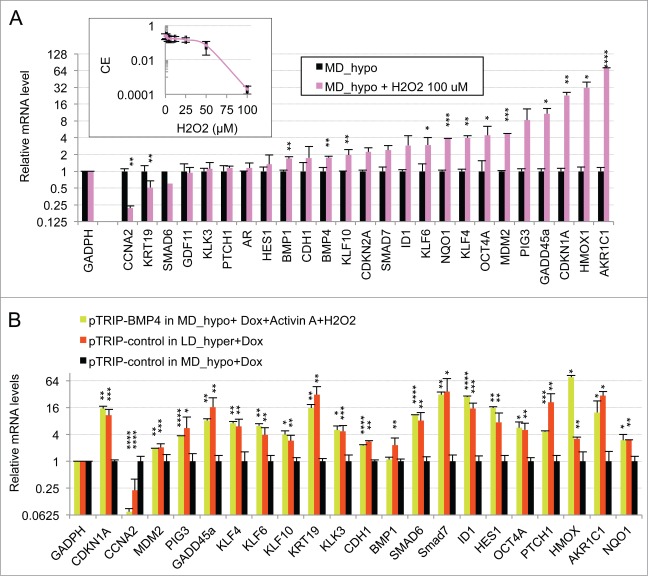
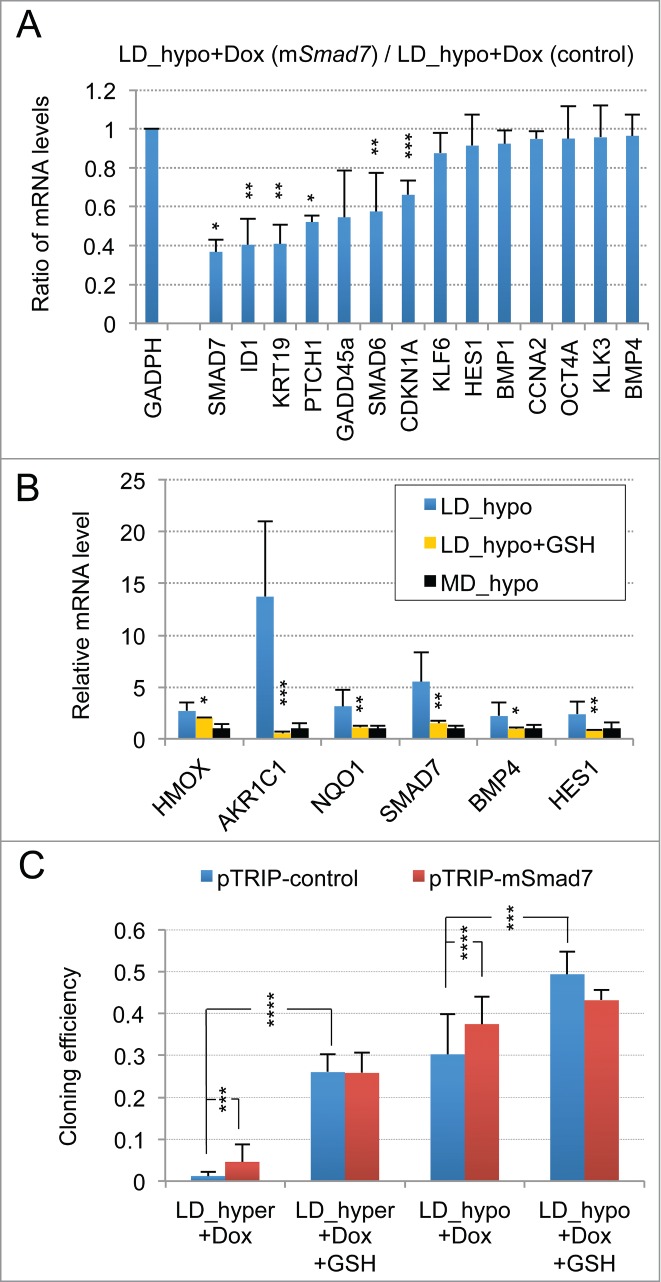
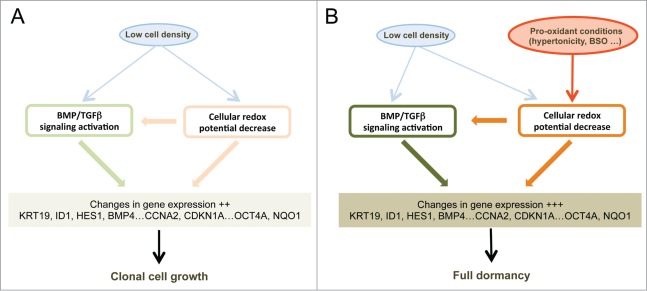
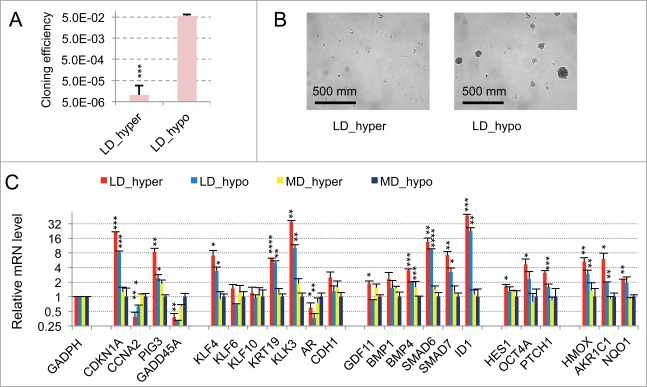
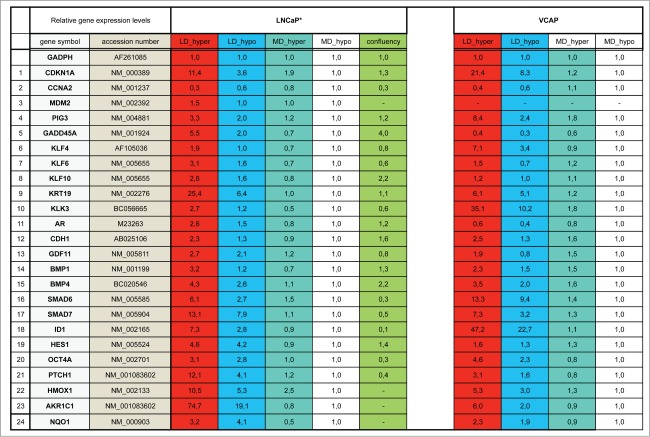
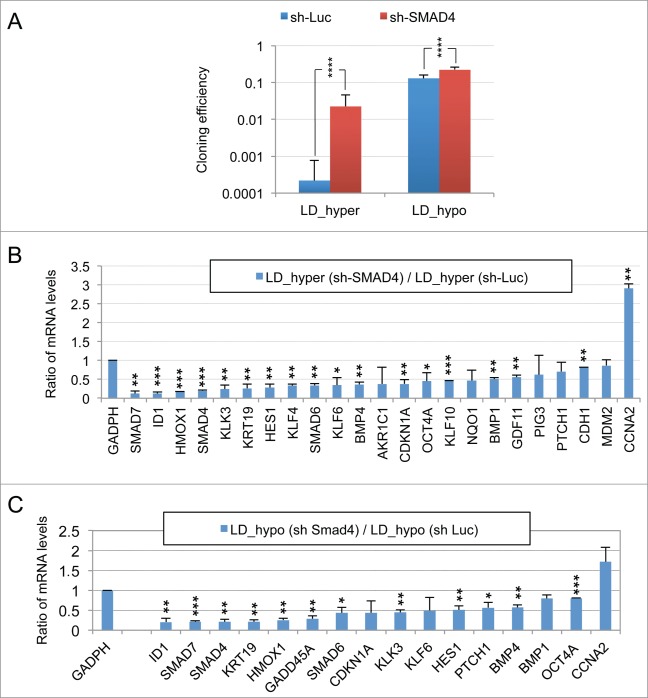
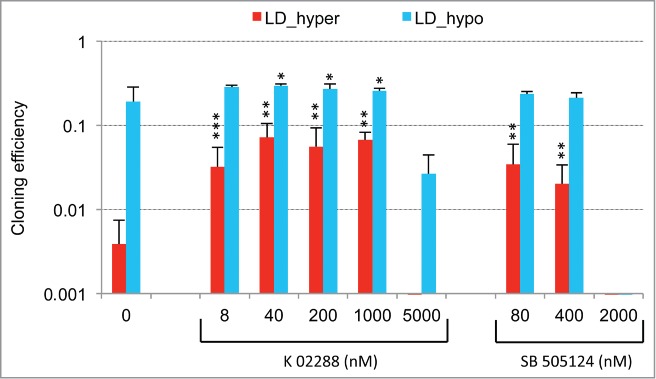
Metastasis involves the dissemination of single or small clumps of cancer cells through blood or lymphatic vessels and their extravasation into distant organs. Despite the strong regulation of metastases development by a cell dormancy phenomenon, the dormant state of cancer cells remains poorly characterized due to the difficulty of in vivo studies. We have recently shown in vitro that clonogenicity of prostate cancer cells is regulated by a dormancy phenomenon that is strongly induced when cells are cultured both at low cell density and in a slightly hypertonic medium. Here, we characterized by RT-qPCR a genetic expression signature of this dormant state which combines the presence of both stemness and differentiation markers. We showed that both TFGβ/BMP signaling and redox imbalance are required for the full induction of this dormancy signature and cell quiescence. Moreover, reconstruction experiments showed that TFGβ/BMP signaling and redox imbalance are sufficient to generate a pattern of genetic expression displaying all characteristic features of the dormancy signature. Finally, we observed that low cell density was sufficient to activate TGFβ/BMP signaling and to generate a slight redox imbalance thus priming cells for dormancy that can be attained with a co-stimulus like hypertonicity, most likely through an increased redox imbalance. The identification of a dual regulation of dormancy provides a framework for the interpretation of previous reports showing a restricted ability of BMP signaling to regulate cancer cell dormancy in vivo and draws attention on the role of oxidative stress in the metastatic process.
Keywords: ARM, Averaging Ratio Method; BSO, buthionine sulfoximine; CE, cloning efficiency; DMEM-FCS, Dulbecco Modified Essential Medium supplemented with 10% Fetal Calf Serum and penicillin-streptomycin; LD_hyper, low cell density condition in DMEM-FCS medium; LD_hypo, low cell density condition in hypotonic medium made by addition of 25% water to DMEM-FCS; MD_hyper, medium cell density condition in DMEM-FCS medium (slightly hypertonic); MD_hypo, medium cell density condition in hypotonic medium made by addition of 25% water to DMEM-FCS; NAM, Normalized Averaging Method; SMAD transcription factor; cell proliferation; cell signaling; prostate cancer; redox regulation.
Figures
References
-
- Aguirre-Ghiso JA, Ossowski L, Rosenbaum SK. Green fluorescent protein tagging of extracellular signal-regulated kinase and p38 pathways reveals novel dynamics of pathway activation during primary and metastatic growth. Cancer Res 2004; 64:7336-45; PMID:15492254; http://dx.doi.org/ 10.1158/0008-5472.CAN-04-0113 - DOI - PubMed
-
- Cameron MD, Schmidt EE, Kerkvliet N, Nadkarni KV, Morris VL, Groom AC, Chambers AF, MacDonald IC. Temporal progression of metastasis in lung: cell survival, dormancy, and location dependence of metastatic inefficiency. Cancer Res 2000; 60:2541-6; PMID:10811137 - PubMed
-
- Goodison S, Kawai K, Hihara J, Jiang P, Yang M, Urquidi V, Hoffman RM, Tarin D. Prolonged dormancy and site-specific growth potential of cancer cells spontaneously disseminated from nonmetastatic breast tumors as revealed by labeling with green fluorescent protein. Clin Cancer Res 2003; 9:3808-14; PMID:14506175 - PubMed
-
- Luzzi KJ, MacDonald IC, Schmidt EE, Kerkvliet N, Morris VL, Chambers AF, Groom AC. Multistep nature of metastatic inefficiency: dormancy of solitary cells after successful extravasation and limited survival of early micrometastases. Am J Pathol 1998; 153:865-73; PMID:9736035; http://dx.doi.org/ 10.1016/S0002-9440(10)65628-3 - DOI - PMC - PubMed
-
- Naumov GN, MacDonald IC, Weinmeister PM, Kerkvliet N, Nadkarni KV, Wilson SM, Morris VL, Groom AC, Chambers AF. Persistence of solitary mammary carcinoma cells in a secondary site: a possible contributor to dormancy. Cancer Res 2002; 62:2162-8; PMID:11929839 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials