

Crystal structure of the V(D)J recombinase RAG1-RAG2
- PMID: 25707801
- PMCID: PMC4342785
- DOI: 10.1038/nature14174
Crystal structure of the V(D)J recombinase RAG1-RAG2
Abstract
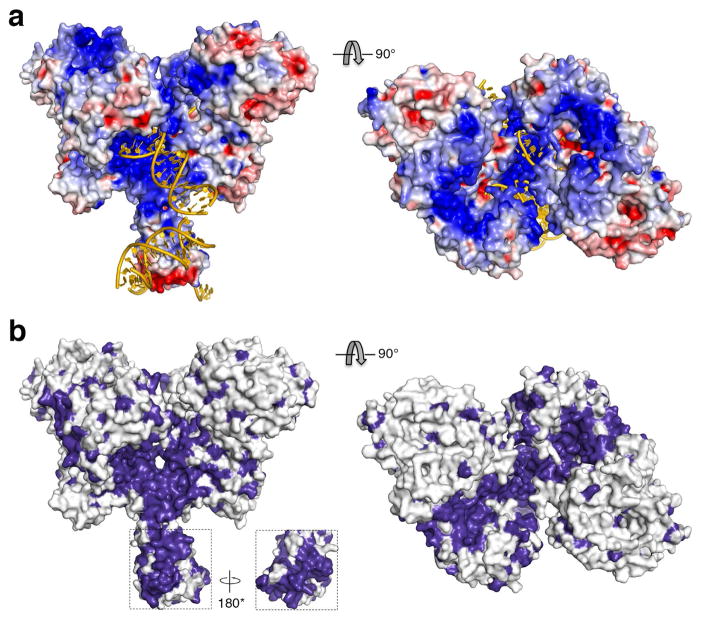
V(D)J recombination in the vertebrate immune system generates a highly diverse population of immunoglobulins and T-cell receptors by combinatorial joining of segments of coding DNA. The RAG1-RAG2 protein complex initiates this site-specific recombination by cutting DNA at specific sites flanking the coding segments. Here we report the crystal structure of the mouse RAG1-RAG2 complex at 3.2 Å resolution. The 230-kilodalton RAG1-RAG2 heterotetramer is 'Y-shaped', with the amino-terminal domains of the two RAG1 chains forming an intertwined stalk. Each RAG1-RAG2 heterodimer composes one arm of the 'Y', with the active site in the middle and RAG2 at its tip. The RAG1-RAG2 structure rationalizes more than 60 mutations identified in immunodeficient patients, as well as a large body of genetic and biochemical data. The architectural similarity between RAG1 and the hairpin-forming transposases Hermes and Tn5 suggests the evolutionary conservation of these DNA rearrangements.
Conflict of interest statement
The authors declare no competing financial interests.
Figures











References
-
- Sakano H, Huppi K, Heinrich G, Tonegawa S. Sequences at the somatic recombination sites of immunoglobulin light-chain genes. Nature. 1979;280:288–294. - PubMed
-
- Lewis SM. The mechanism of V(D)J joining: lessons from molecular, immunological, and comparative analyses. Adv Immunol. 1994;56:27–150. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
