

RNA degradation in antiviral immunity and autoimmunity
- PMID: 25709093
- PMCID: PMC4358841
- DOI: 10.1016/j.it.2015.02.001
RNA degradation in antiviral immunity and autoimmunity
Abstract
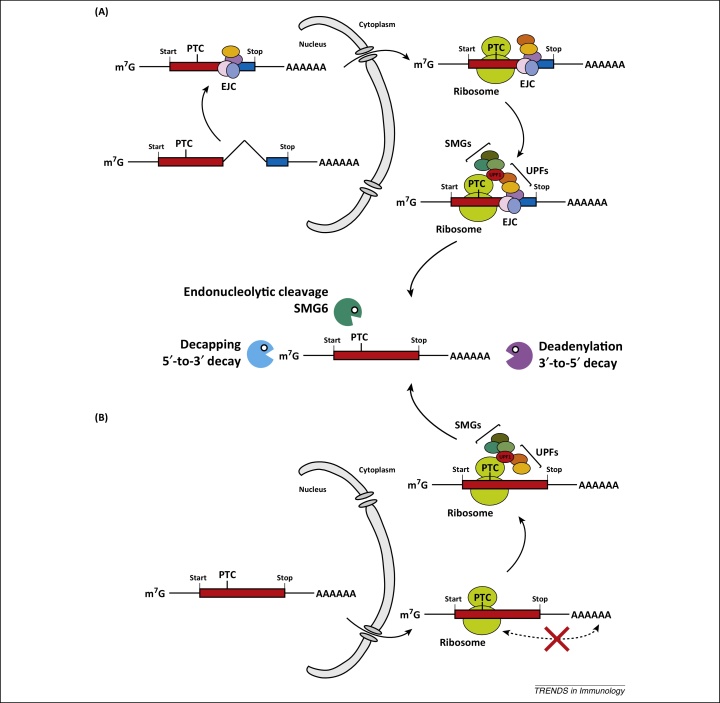
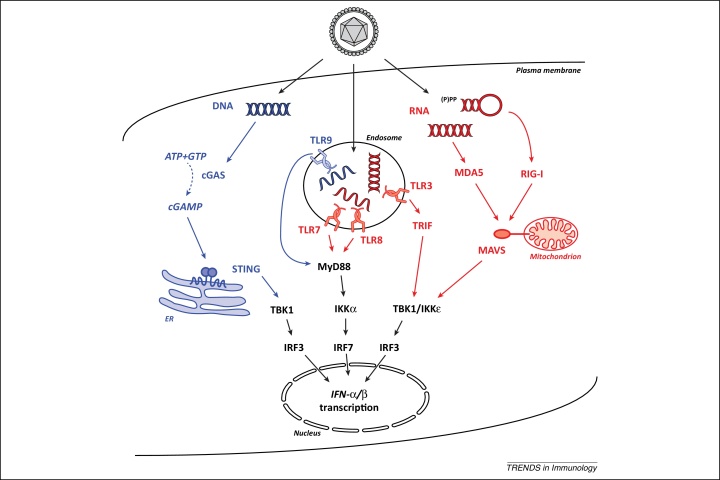
Post-transcriptional control determines the fate of cellular RNA molecules. Nonsense-mediated decay (NMD) provides quality control of mRNA, targeting faulty cellular transcripts for degradation by multiple nucleases including the RNA exosome. Recent findings have revealed a role for NMD in targeting viral RNA molecules, thereby restricting virus infection. Interestingly, NMD is also linked to immune responses at another level: mutations affecting the NMD or RNA exosome machineries cause chronic activation of defence programmes, resulting in autoimmune phenotypes. Here we place these observations in the context of other links between innate antiviral immunity and type I interferon mediated disease and examine two models: one in which expression or function of pathogen sensors is perturbed and one wherein host-derived RNA molecules with a propensity to activate such sensors accumulate.
Keywords: Aicardi-Goutières syndrome; MDA5; RNA exosome; nonsense-mediated decay; pattern-recognition receptor; type I interferon.
Copyright © 2015 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
References
-
- Balistreri G. The host nonsense-mediated mRNA decay pathway restricts Mammalian RNA virus replication. Cell Host Microbe. 2014;16:403–411. - PubMed
-
- Gloggnitzer J. Nonsense-mediated mRNA decay modulates immune receptor levels to regulate plant antibacterial defense. Cell Host Microbe. 2014;16:376–390. - PubMed
-
- Behm-Ansmant I. mRNA quality control: an ancient machinery recognizes and degrades mRNAs with nonsense codons. FEBS Lett. 2007;581:2845–2853. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
