

Methylglyoxal, the dark side of glycolysis
- PMID: 25709564
- PMCID: PMC4321437
- DOI: 10.3389/fnins.2015.00023
Methylglyoxal, the dark side of glycolysis
Abstract
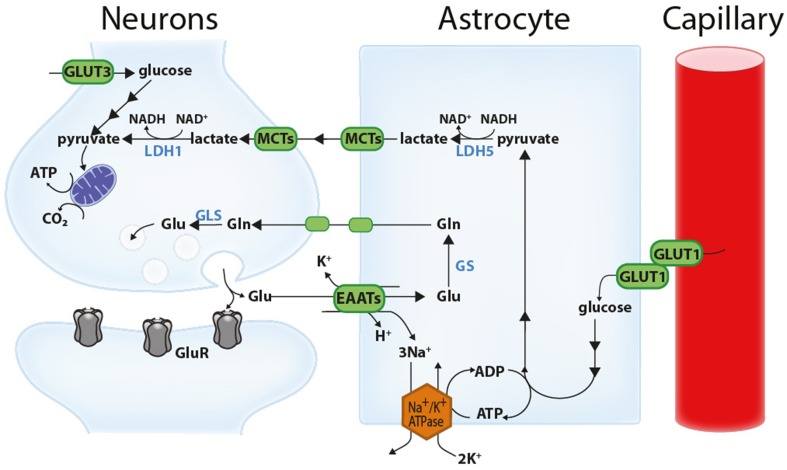
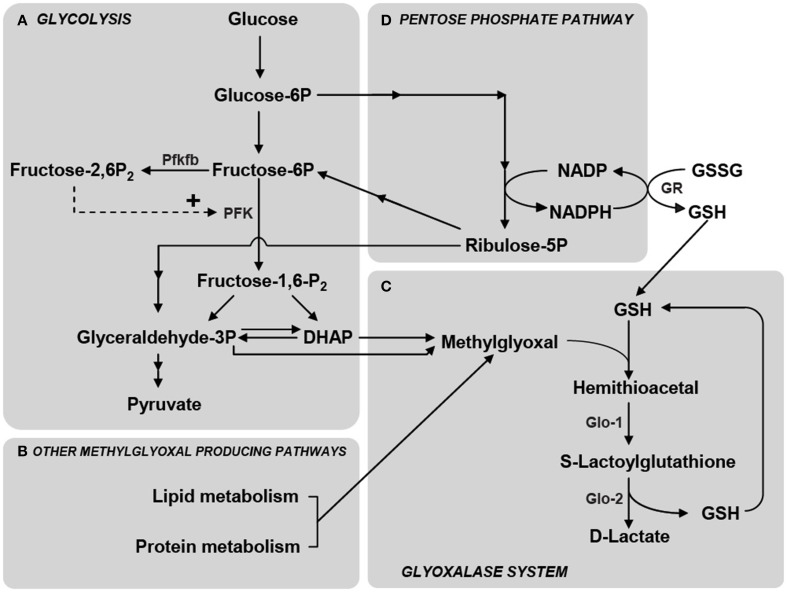
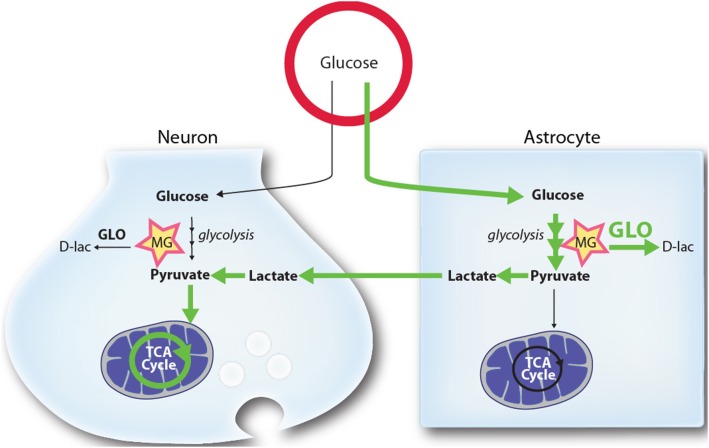
Glucose is the main energy substrate for the brain. There is now extensive evidence indicating that the metabolic profile of neural cells with regard to glucose utilization and glycolysis rate is not homogenous, with a marked propensity for glycolytic glucose processing in astrocytes compared to neurons. Methylglyoxal, a highly reactive dicarbonyl compound, is inevitably formed as a by-product of glycolysis. Methylglyoxal is a major cell-permeant precursor of advanced glycation end-products (AGEs), which are associated with several pathologies including diabetes, aging and neurodegenerative diseases. In normal situations, cells are protected against methylglyoxal toxicity by different mechanisms and in particular the glyoxalase system, which represents the most important pathway for the detoxification of methylglyoxal. While the neurotoxic effects of methylglyoxal and AGEs are well characterized, our understanding the glyoxalase system in the brain is more scattered. Considering the high energy requirements (i.e., glucose) of the brain, one should expect that the cerebral glyoxalase system is adequately fitted to handle methylglyoxal toxicity. This review focuses on our actual knowledge on the cellular aspects of the glyoxalase system in brain cells, in particular with regard to its activity in astrocytes and neurons. A main emerging concept is that these two neural cell types have different and energetically adapted glyoxalase defense mechanisms which may serve as protective mechanism against methylglyoxal-induced cellular damage.
Keywords: advanced-glycation end-products (AGEs); astrocyte; glutathione; methylglyoxal; neuron; triosephosphate.
Figures
References
-
- Ahmed N., Battah S., Karachalias N., Babaei-Jadidi R., Horanyi M., Baroti K., et al. (2003a). Increased formation of methylglyoxal and protein glycation, oxidation and nitrosation in triosephosphate isomerase deficiency. Biochim. Biophys. Acta 1639, 121–132. 10.1016/j.bbadis.2003.08.002 - DOI - PubMed
-
- Allaman I. (2009). Glial glycogen metabolism, in Encyclopedia of Neuroscience, ed Squire L. R. (Oxford: Academic Press; ), 811–818.
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources