

Licochalcone A, a polyphenol present in licorice, suppresses UV-induced COX-2 expression by targeting PI3K, MEK1, and B-Raf
- PMID: 25710724
- PMCID: PMC4394430
- DOI: 10.3390/ijms16034453
Licochalcone A, a polyphenol present in licorice, suppresses UV-induced COX-2 expression by targeting PI3K, MEK1, and B-Raf
Abstract
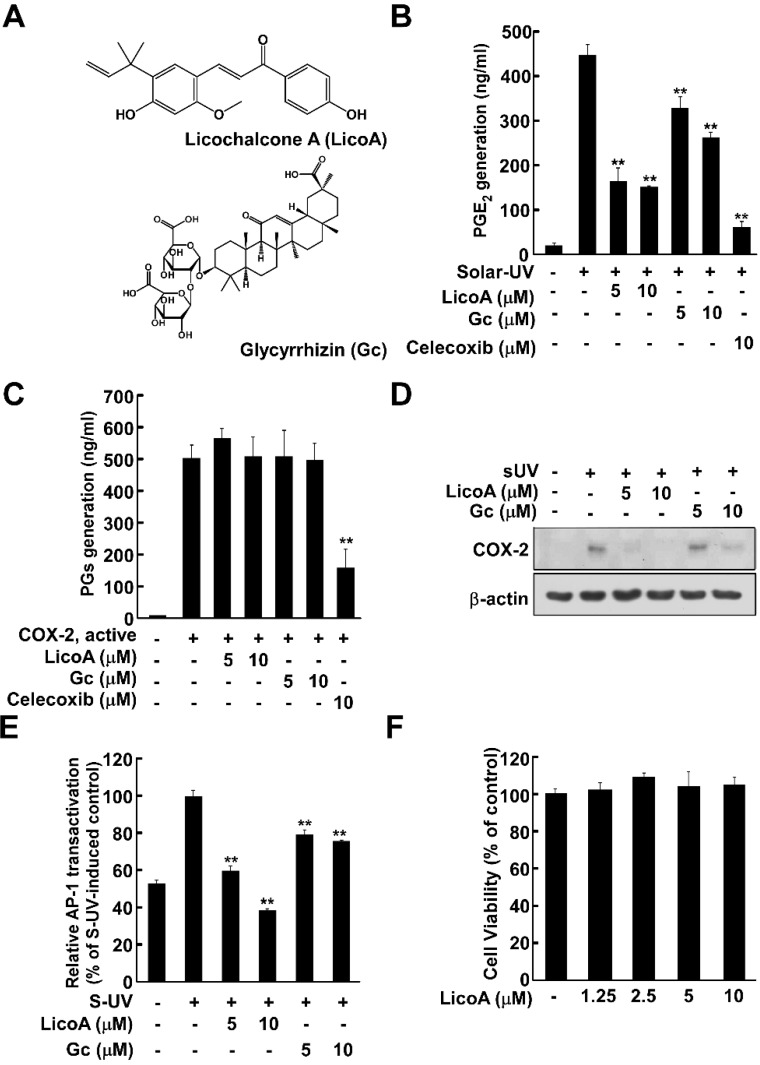
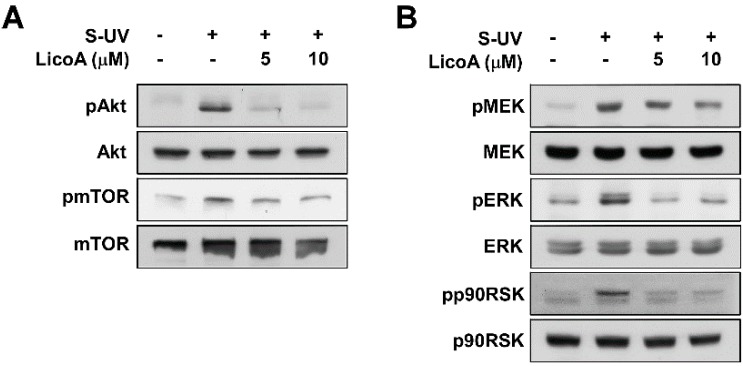
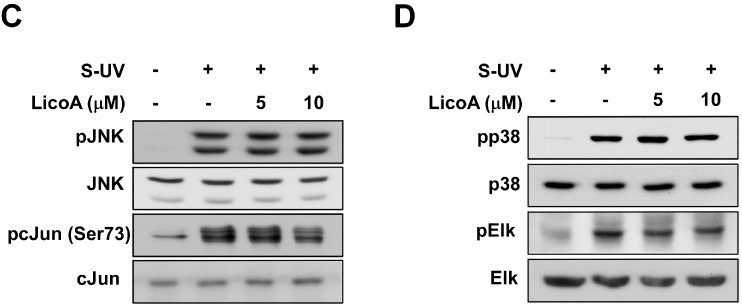
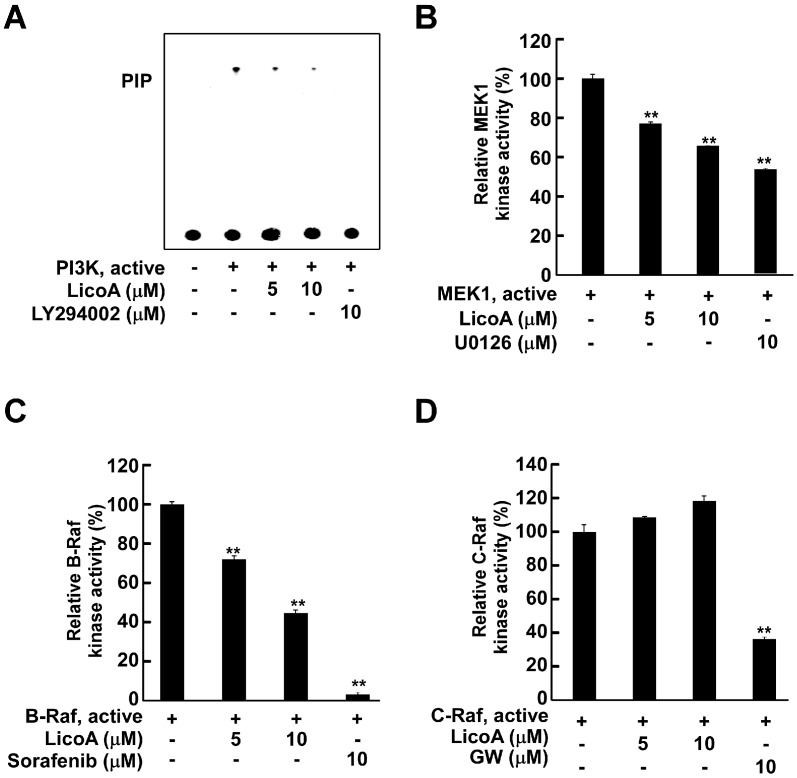
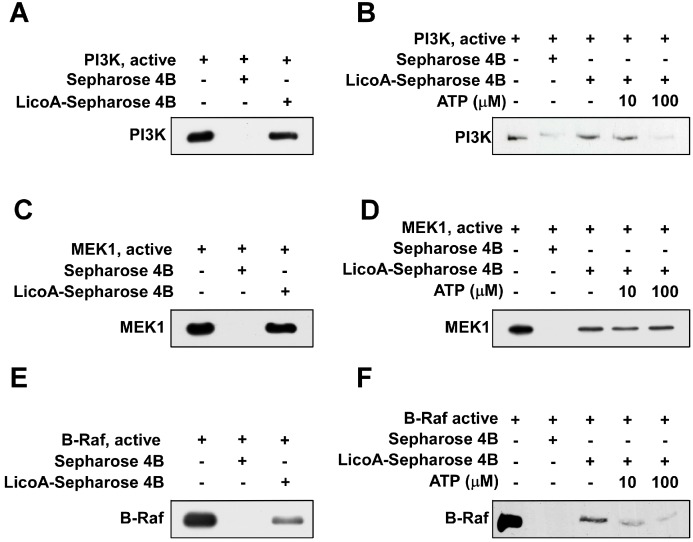
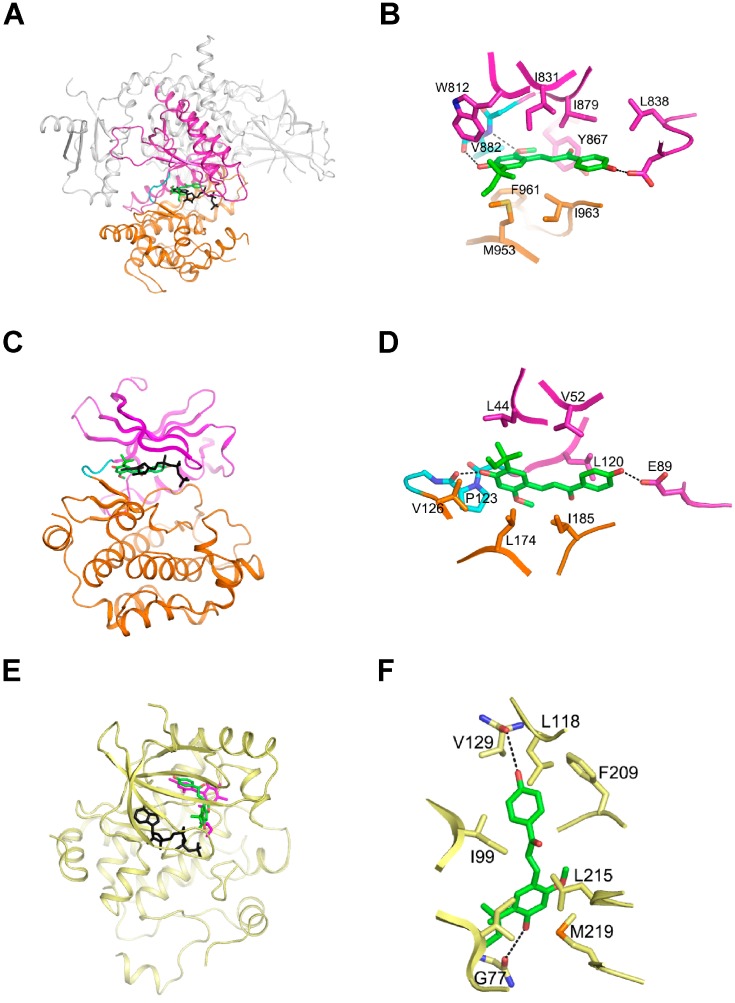
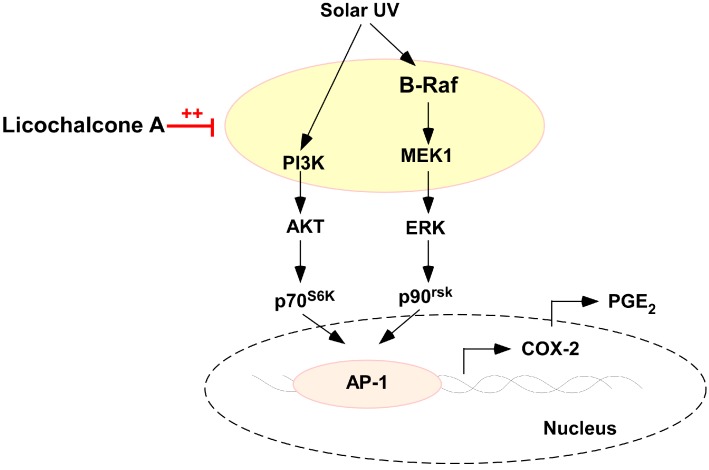
Licorice is a traditional botanical medicine, and has historically been commonly prescribed in Asia to treat various diseases. Glycyrrhizin (Gc), a triterpene compound, is the most abundant phytochemical constituent of licorice. However, high intake or long-term consumption of Gc has been associated with a number of side effects, including hypertension. However, the presence of alternative bioactive compounds in licorice with anti-carcinogenic effects has long been suspected. Licochalcone A (LicoA) is a prominent member of the chalcone family and can be isolated from licorice root. To date, there have been no reported studies on the suppressive effect of LicoA against solar ultraviolet (sUV)-induced cyclooxygenase (COX)-2 expression and the potential molecular mechanisms involved. Here, we show that LicoA, a major chalcone compound of licorice, effectively inhibits sUV-induced COX-2 expression and prostaglandin E2 PGE2 generation through the inhibition of activator protein 1 AP-1 transcriptional activity, with an effect that is notably more potent than Gc. Western blotting analysis shows that LicoA suppresses sUV-induced phosphorylation of Akt/ mammalian target of rapamycin (mTOR) and extracellular signal-regulated kinases (ERK)1/2/p90 ribosomal protein S6 kinase (RSK) in HaCaT cells. Moreover, LicoA directly suppresses the activity of phosphoinositide 3-kinase (PI3K), mitogen-activated protein kinase kinase (MEK)1, and B-Raf, but not Raf-1 in cell-free assays, indicating that PI3K, MEK1, and B-Raf are direct molecular targets of LicoA. We also found that LicoA binds to PI3K and B-Raf in an ATP-competitive manner, although LicoA does not appear to compete with ATP for binding with MEK1. Collectively, these results provide insight into the biological action of LicoA, which may have potential for development as a skin cancer chemopreventive agent.
Figures
References
-
- Kuwajima H., Taneda Y., Chen W.Z., Kawanishi T., Hori K., Taniyama T., Kobayashi M., Ren J., Kitagawa I. Variation of chemical constituents in processed licorice roots: Quantitative determination of saponin and flavonoid constituents in bark removed and roasted licorice roots. Yakugaku Zasshi. 1999;119:945–955. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
