

Ras-mediated modulation of pyruvate dehydrogenase activity regulates mitochondrial reserve capacity and contributes to glioblastoma tumorigenesis
- PMID: 25712957
- PMCID: PMC4588752
- DOI: 10.1093/neuonc/nou369
Ras-mediated modulation of pyruvate dehydrogenase activity regulates mitochondrial reserve capacity and contributes to glioblastoma tumorigenesis
Erratum in
-
Corrigendum to: "Ras-mediated modulation of pyruvate dehydrogenase activity regulates mitochondrial reserve capacity and contributes to glioblastoma tumorigenesis".Neuro Oncol. 2024 Dec 5;26(12):2395. doi: 10.1093/neuonc/noae156. Neuro Oncol. 2024. PMID: 39221889 Free PMC article. No abstract available.
Abstract
Background: Even though altered metabolism representing a hallmark of cancer was proposed nearly a century ago, recent technological advances have allowed investigators to continue uncovering a previously unrecognized complexity of metabolic programs that drive tumorigenesis beyond that of aerobic glycolysis.
Methods: The bioenergetic state of a diverse panel of glioblastoma models, including isogenic lines derived from a genetically engineered adult astrocytic mouse model and patient-derived glioblastoma stem cells, was determined at baseline and in stressed conditions. Mechanisms contributing to the discovered metabolic phenotypes were determined through molecular and chemical perturbation, and their biological consequences were evaluated in vivo and in patient samples.
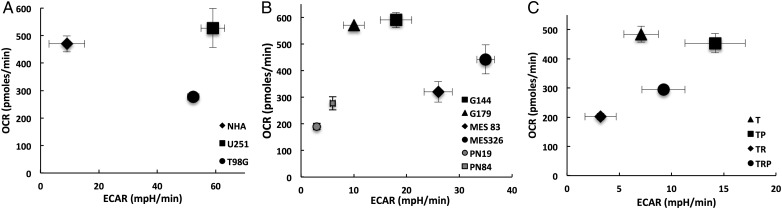
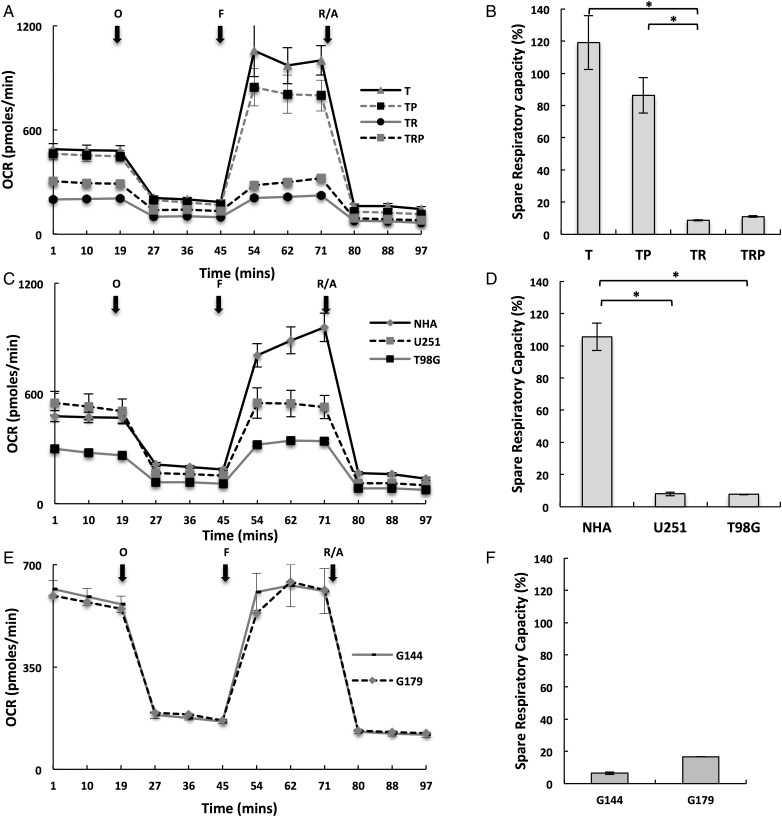
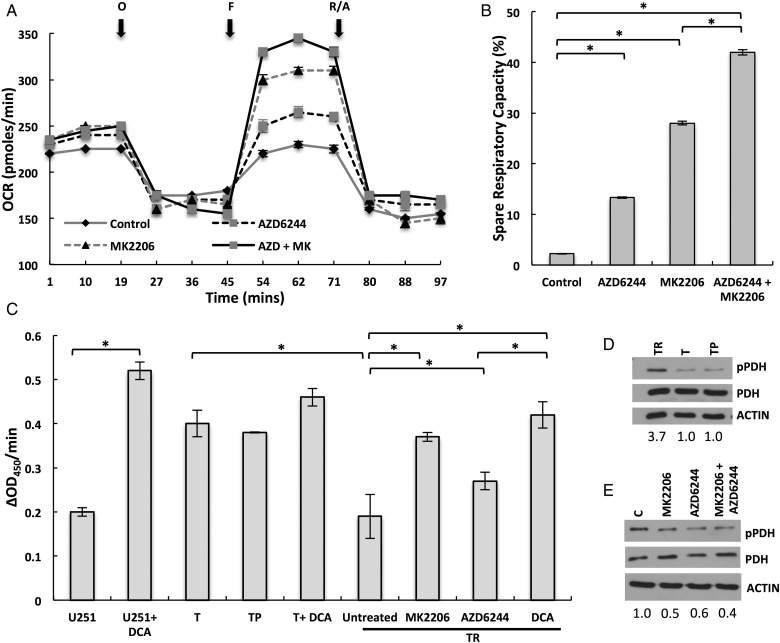
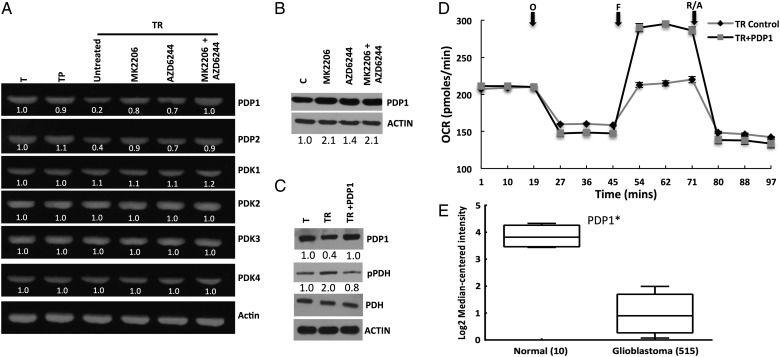
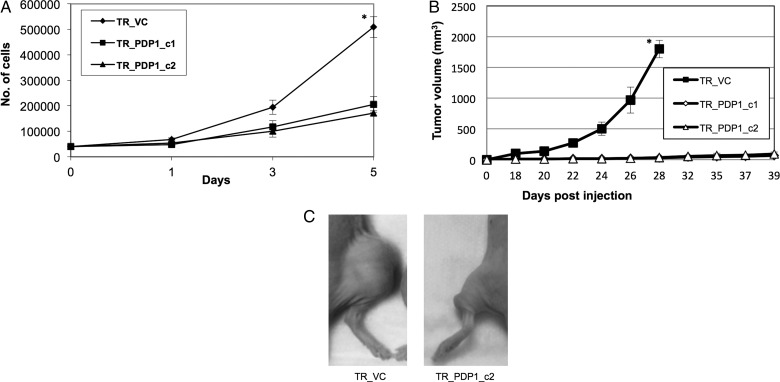
Results: Attenuated mitochondrial reserve capacity was identified as a common metabolic phenotype in glioblastoma lines. This phenotype was linked mechanistically with the capacity of Ras-mediated signaling to inhibit pyruvate dehydrogenase (PDH) activity through downregulation of PDH phosphatase (PDP) expression. PDP1 repression was validated clinically in patient-derived samples, suggesting that aberrant cellular signaling typical of glioblastoma actively modulates PDH activity. This phenotype was reversed through both chemical and molecular perturbation. Restoration of PDH activity through stable expression of PDP1-impaired tumorigenic potential.
Conclusions: These findings support the central role that PDH regulation plays as a downstream consequence of aberrant signaling associated with gliomagenesis and the scientific rationale to continue to develop and test clinical strategies designed to activate PDH as a form of anticancer therapy in glioblastoma.
Keywords: Ras; glioblastoma; pyruvate dehydrogenase; spare respiratory capacity; tumor metabolism.
© The Author(s) 2015. Published by Oxford University Press on behalf of the Society for Neuro-Oncology. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
References
-
- Warburg O. On respiratory impairment in cancer cells. Science. 1956;124(3215):269–270. - PubMed
-
- Warburg O, Posener K, Negelein E. Uber den Stoffwechsel der Carcinomzelle. Biochem Zeitschr. 1924;152:309–344.
-
- Moreno-Sanchez R, Rodriguez-Enriquez S, Marin-Hernandez A, Saavedra E. Energy metabolism in tumor cells. FEBS J. 2007;274(6):1393–1418. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
