

Family 46 Carbohydrate-binding Modules Contribute to the Enzymatic Hydrolysis of Xyloglucan and β-1,3-1,4-Glucans through Distinct Mechanisms
- PMID: 25713075
- PMCID: PMC4409224
- DOI: 10.1074/jbc.M115.637827
Family 46 Carbohydrate-binding Modules Contribute to the Enzymatic Hydrolysis of Xyloglucan and β-1,3-1,4-Glucans through Distinct Mechanisms
Abstract
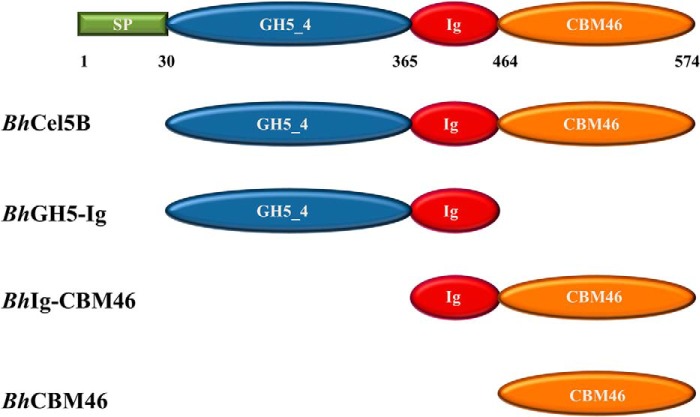
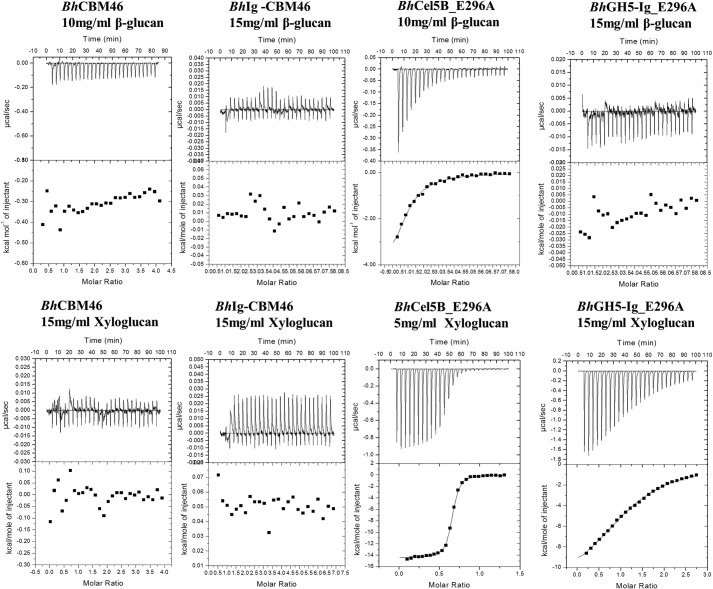
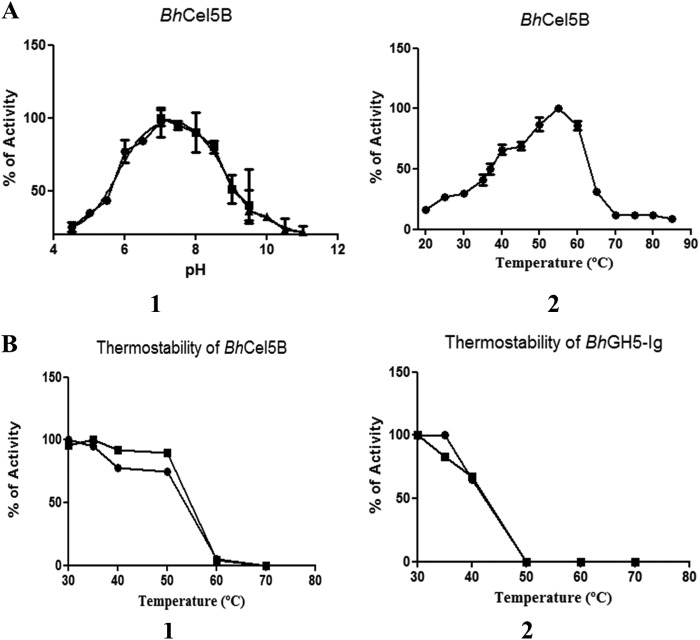
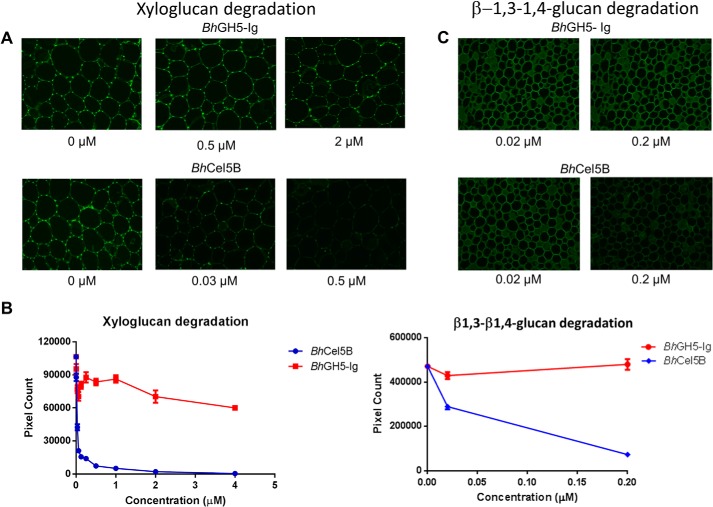
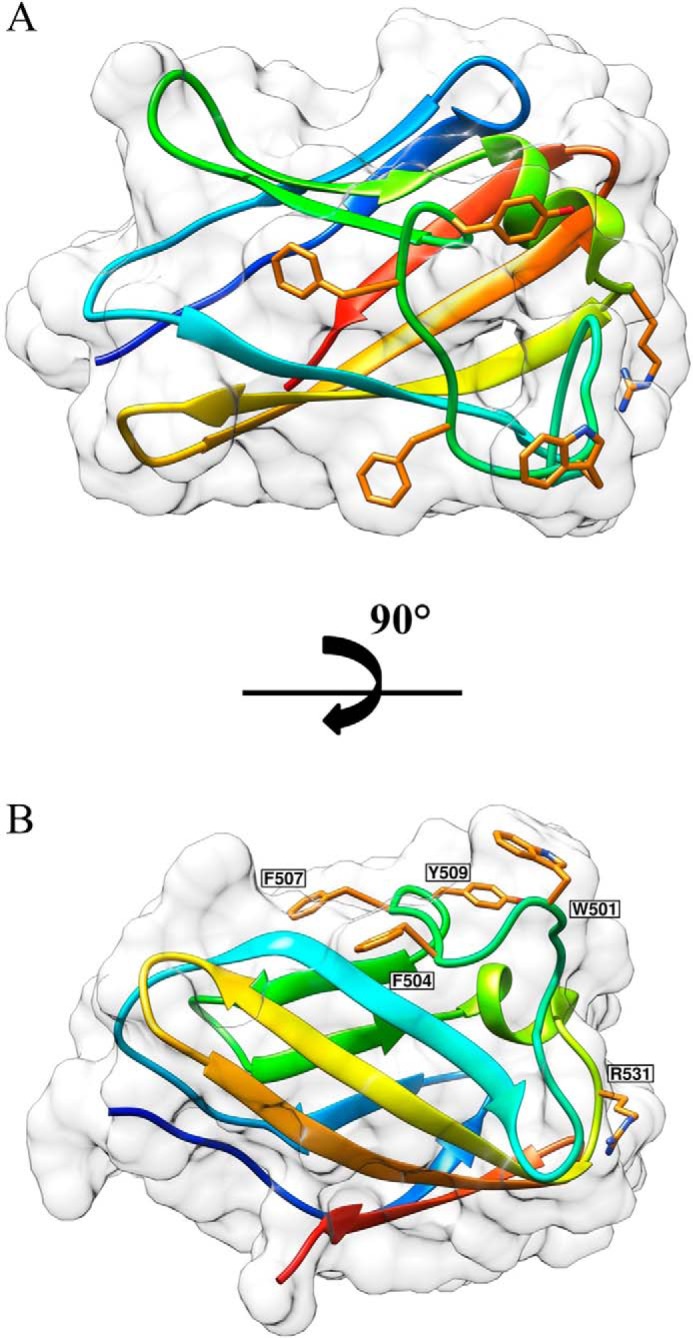
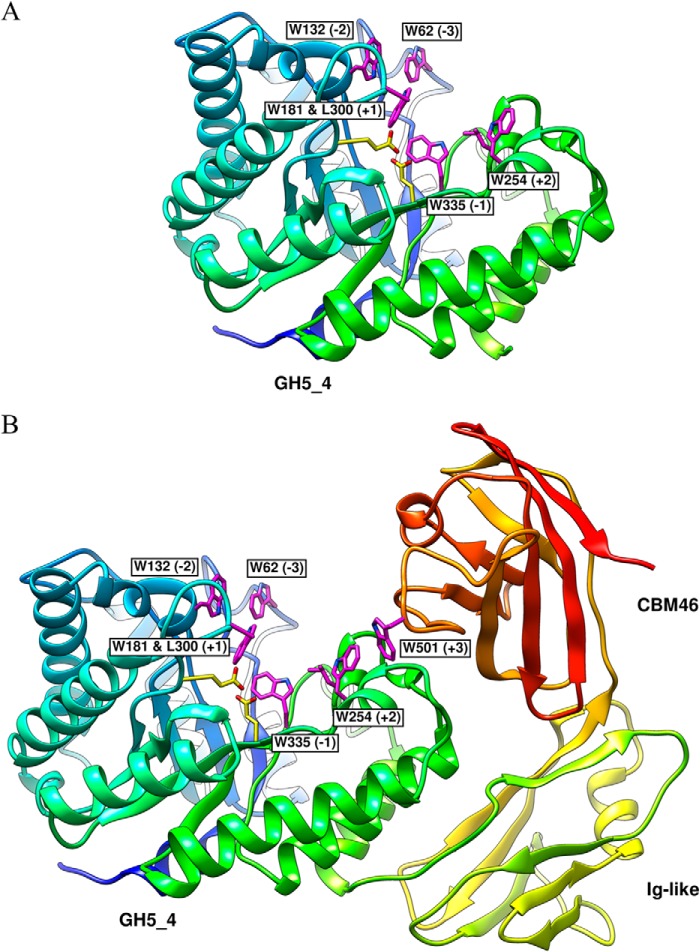
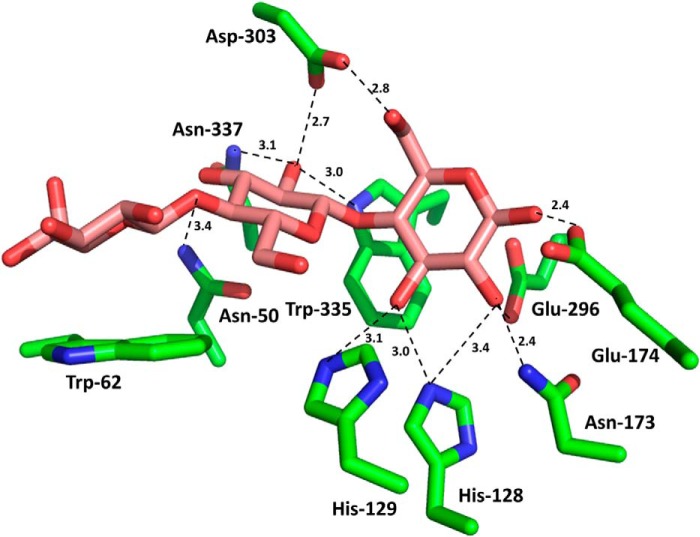
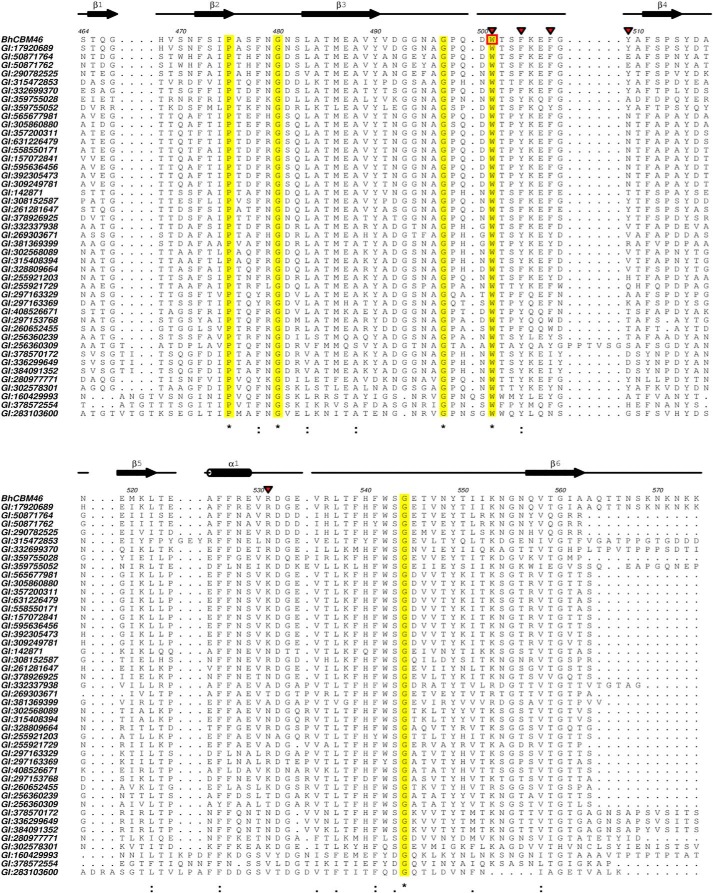
Structural carbohydrates comprise an extraordinary source of energy that remains poorly utilized by the biofuel sector as enzymes have restricted access to their substrates within the intricacy of plant cell walls. Carbohydrate active enzymes (CAZYmes) that target recalcitrant polysaccharides are modular enzymes containing noncatalytic carbohydrate-binding modules (CBMs) that direct enzymes to their cognate substrate, thus potentiating catalysis. In general, CBMs are functionally and structurally autonomous from their associated catalytic domains from which they are separated through flexible linker sequences. Here, we show that a C-terminal CBM46 derived from BhCel5B, a Bacillus halodurans endoglucanase, does not interact with β-glucans independently but, uniquely, acts cooperatively with the catalytic domain of the enzyme in substrate recognition. The structure of BhCBM46 revealed a β-sandwich fold that abuts onto the region of the substrate binding cleft upstream of the active site. BhCBM46 as a discrete entity is unable to bind to β-glucans. Removal of BhCBM46 from BhCel5B, however, abrogates binding to β-1,3-1,4-glucans while substantially decreasing the affinity for decorated β-1,4-glucan homopolymers such as xyloglucan. The CBM46 was shown to contribute to xyloglucan hydrolysis only in the context of intact plant cell walls, but it potentiates enzymatic activity against purified β-1,3-1,4-glucans in solution or within the cell wall. This report reveals the mechanism by which a CBM can promote enzyme activity through direct interaction with the substrate or by targeting regions of the plant cell wall where the target glucan is abundant.
Keywords: Carbohydrate-binding Protein; Cellulase; Cellulose; Plant Cell Wall; Protein Structure.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Himmel M. E., Bayer E. A. (2009) Lignocellulose conversion to biofuels: current challenges, global perspectives. Curr. Opin Biotechnol. 20, 316–317 - PubMed
-
- Himmel M. E., Ding S. Y., Johnson D. K., Adney W. S., Nimlos M. R., Brady J. W., Foust T. D. (2007) Biomass recalcitrance: engineering plants and enzymes for biofuels production. Science 315, 804–807 - PubMed
-
- Mohnen D. (2008) Pectin structure and biosynthesis. Curr. Opin Plant Biol. 11, 266–277 - PubMed
-
- Minic Z., Jouanin L. (2006) Plant glycoside hydrolases involved in cell wall polysaccharide degradation. Plant Physiol. Biochem. 44, 435–449 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
