

Modeling autosomal recessive cutis laxa type 1C in mice reveals distinct functions for Ltbp-4 isoforms
- PMID: 25713297
- PMCID: PMC4381339
- DOI: 10.1242/dmm.018960
Modeling autosomal recessive cutis laxa type 1C in mice reveals distinct functions for Ltbp-4 isoforms
Abstract
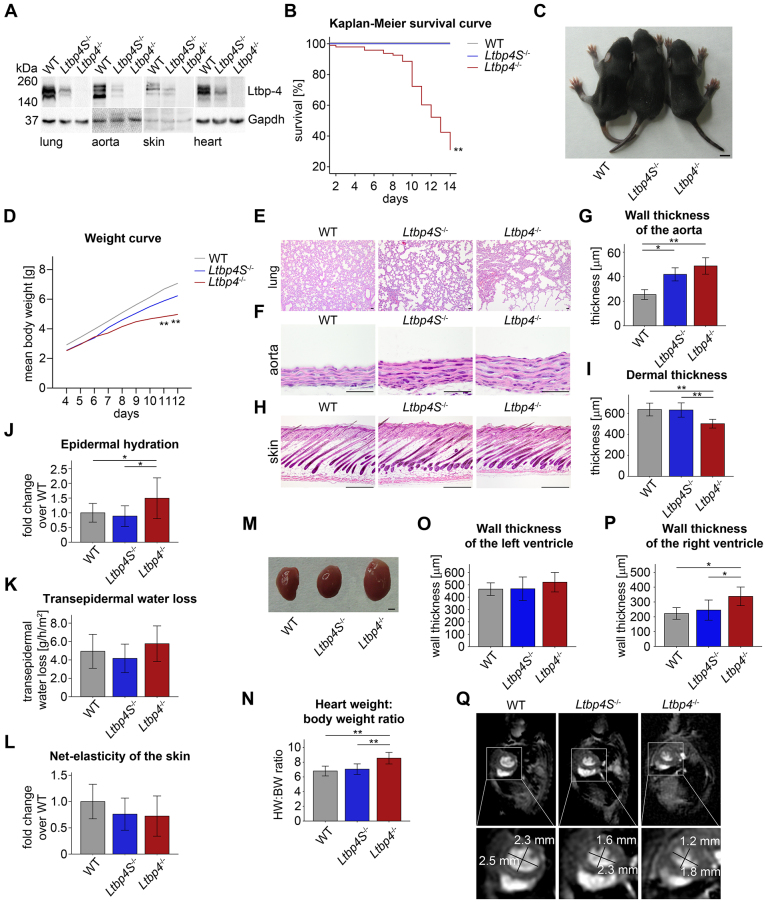
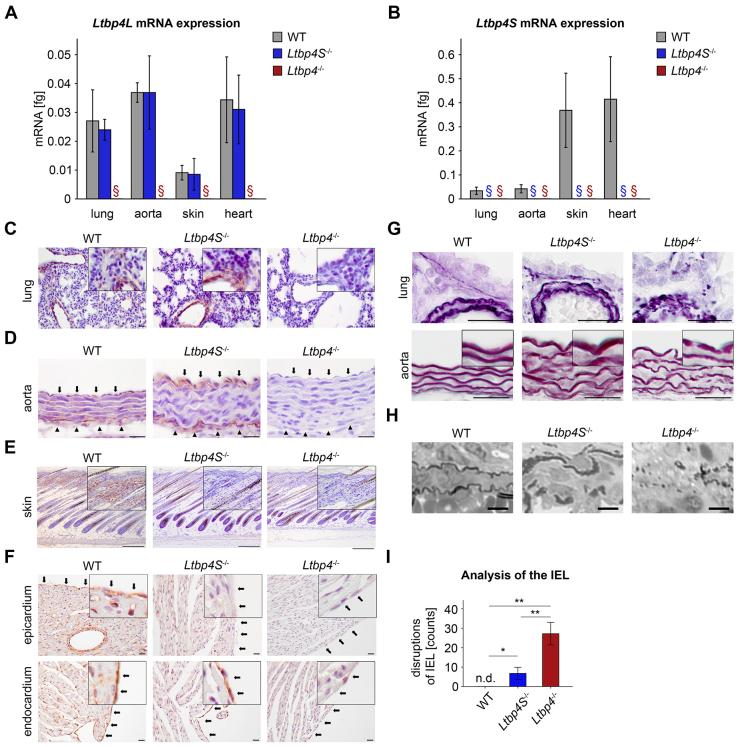
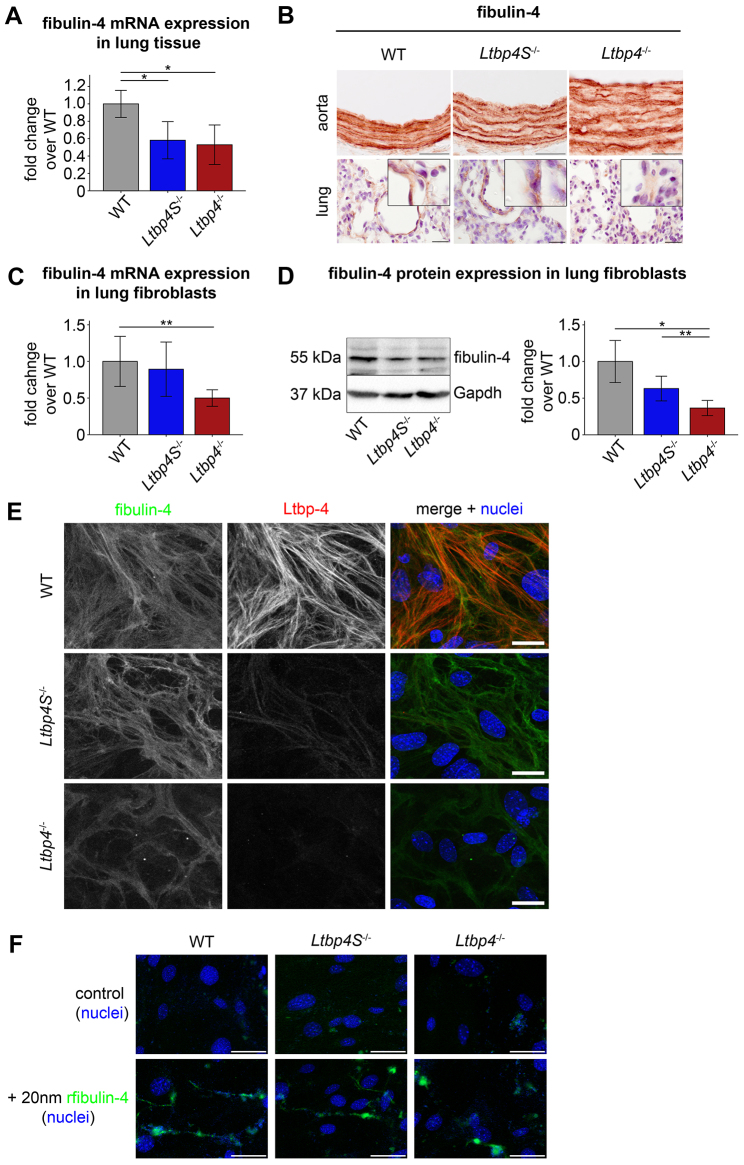
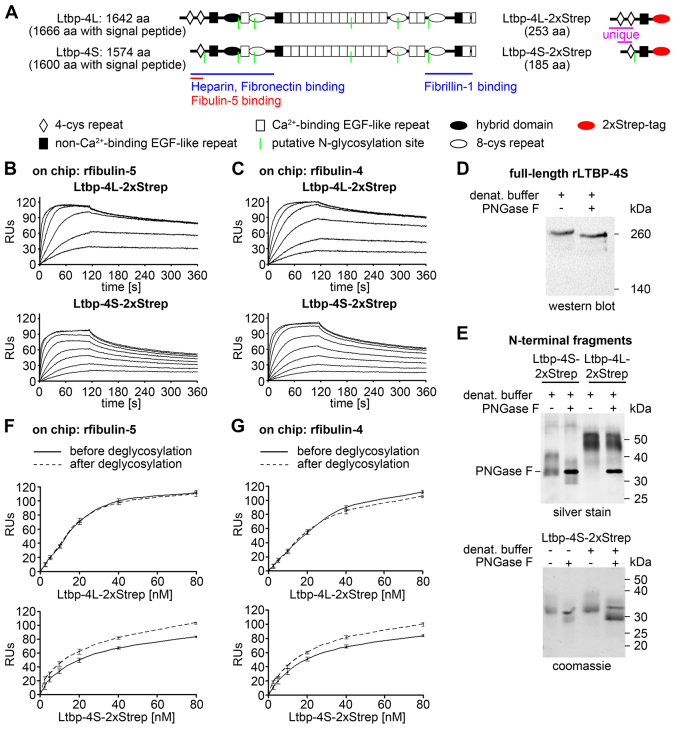
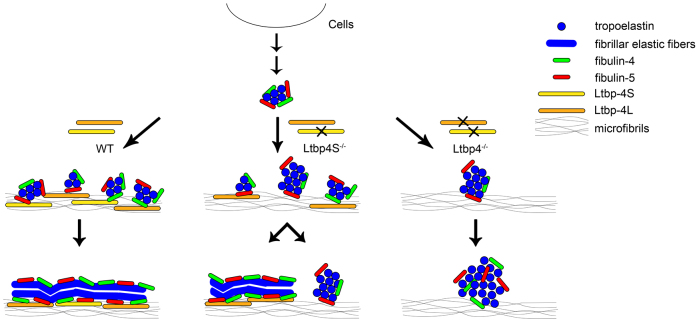
Recent studies have revealed an important role for LTBP-4 in elastogenesis. Its mutational inactivation in humans causes autosomal recessive cutis laxa type 1C (ARCL1C), which is a severe disorder caused by defects of the elastic fiber network. Although the human gene involved in ARCL1C has been discovered based on similar elastic fiber abnormalities exhibited by mice lacking the short Ltbp-4 isoform (Ltbp4S(-/-)), the murine phenotype does not replicate ARCL1C. We therefore inactivated both Ltbp-4 isoforms in the mouse germline to model ARCL1C. Comparative analysis of Ltbp4S(-/-) and Ltbp4-null (Ltbp4(-/-)) mice identified Ltbp-4L as an important factor for elastogenesis and postnatal survival, and showed that it has distinct tissue expression patterns and specific molecular functions. We identified fibulin-4 as a previously unknown interaction partner of both Ltbp-4 isoforms and demonstrated that at least Ltbp-4L expression is essential for incorporation of fibulin-4 into the extracellular matrix (ECM). Overall, our results contribute to the current understanding of elastogenesis and provide an animal model of ARCL1C.
Keywords: ARCL1C; Autosomal recessive cutis laxa type 1C; ECM; Elastogenesis; Extracellular matrix; Fibulin-4; Fibulin-5; Latent transforming growth factor β-binding protein 4; Ltbp-4; Ltbp-4L; Ltbp-4S.
© 2015. Published by The Company of Biologists Ltd.
Figures
References
-
- Abe M., Harpel J. G., Metz C. N., Nunes I., Loskutoff D. J., Rifkin D. B. (1994). An assay for transforming growth factor-beta using cells transfected with a plasminogen activator inhibitor-1 promoter-luciferase construct. Anal. Biochem. 216, 276–284. - PubMed
-
- Annes J., Vassallo M., Munger J. S., Rifkin D. B. (2004). A genetic screen to identify latent transforming growth factor beta activators. Anal. Biochem. 327, 45–54. - PubMed
-
- Bunck A. C., Engelen M. A., Schnackenburg B., Furkert J., Bremer C., Heindel W., Stypmann J., Maintz D. (2009). Feasibility of functional cardiac MR imaging in mice using a clinical 3 Tesla whole body scanner. Invest. Radiol. 44, 749–756. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
