

Nuclear p120-catenin regulates the anoikis resistance of mouse lobular breast cancer cells through Kaiso-dependent Wnt11 expression
- PMID: 25713299
- PMCID: PMC4381336
- DOI: 10.1242/dmm.018648
Nuclear p120-catenin regulates the anoikis resistance of mouse lobular breast cancer cells through Kaiso-dependent Wnt11 expression
Abstract
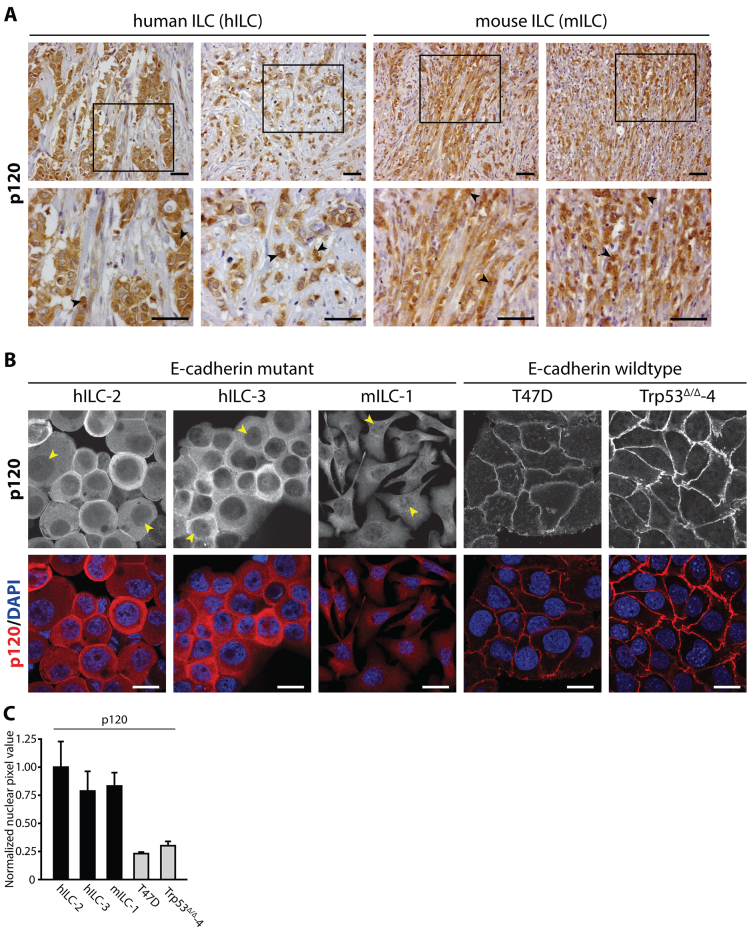
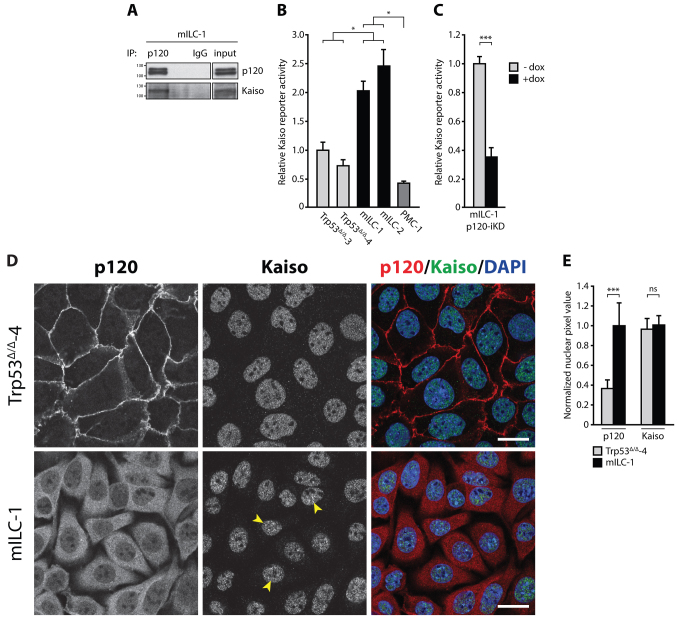
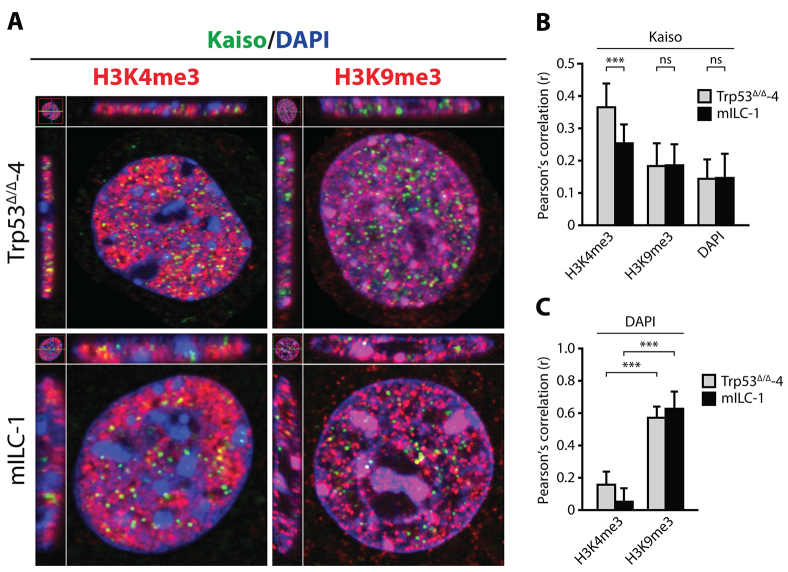
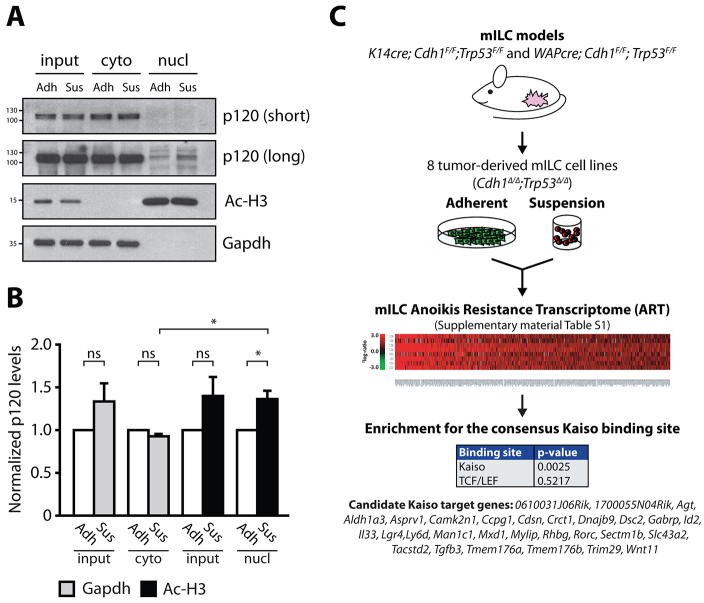
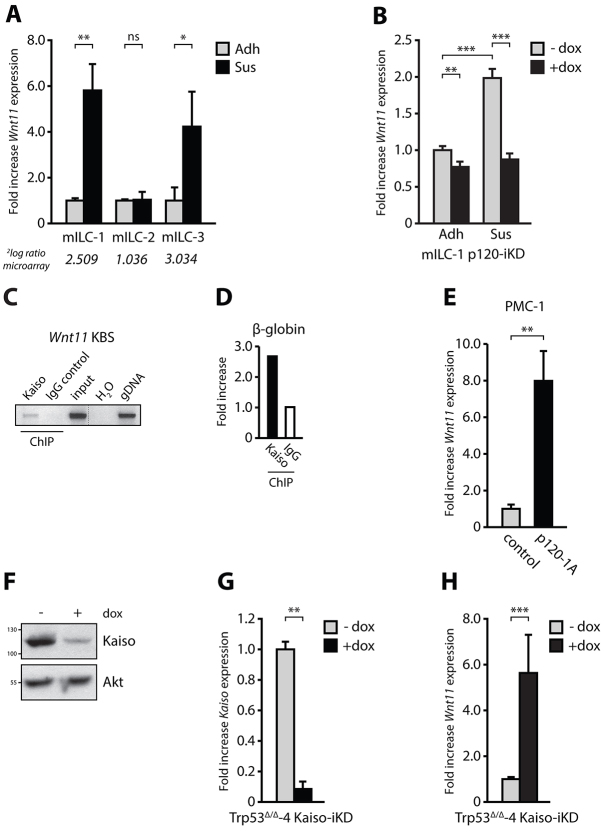
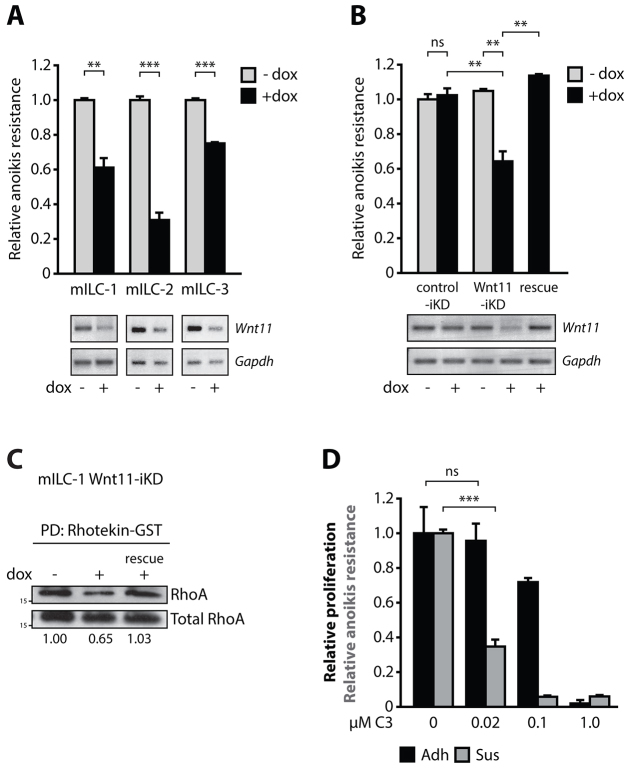
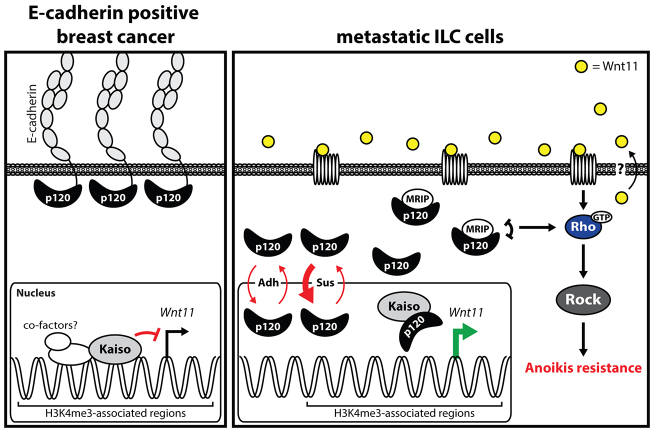
E-cadherin inactivation underpins the progression of invasive lobular breast carcinoma (ILC). In ILC, p120-catenin (p120) translocates to the cytosol where it controls anchorage independence through the Rho-Rock signaling pathway, a key mechanism driving tumor growth and metastasis. We now demonstrate that anchorage-independent ILC cells show an increase in nuclear p120, which results in relief of transcriptional repression by Kaiso. To identify the Kaiso target genes that control anchorage independence we performed genome-wide mRNA profiling on anoikis-resistant mouse ILC cells, and identified 29 candidate target genes, including the established Kaiso target Wnt11. Our data indicate that anchorage-independent upregulation of Wnt11 in ILC cells is controlled by nuclear p120 through inhibition of Kaiso-mediated transcriptional repression. Finally, we show that Wnt11 promotes activation of RhoA, which causes ILC anoikis resistance. Our findings thereby establish a mechanistic link between E-cadherin loss and subsequent control of Rho-driven anoikis resistance through p120- and Kaiso-dependent expression of Wnt11.
Keywords: Anoikis resistance; Breast cancer metastasis; Kaiso; p120-catenin.
© 2015. Published by The Company of Biologists Ltd.
Figures
References
-
- Behrens J., von Kries J. P., Kühl M., Bruhn L., Wedlich D., Grosschedl R., Birchmeier W. (1996). Functional interaction of beta-catenin with the transcription factor LEF-1. Nature 382, 638–642. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
