

Histone titration against the genome sets the DNA-to-cytoplasm threshold for the Xenopus midblastula transition
- PMID: 25713373
- PMCID: PMC4364222
- DOI: 10.1073/pnas.1413990112
Histone titration against the genome sets the DNA-to-cytoplasm threshold for the Xenopus midblastula transition
Abstract
During early development, animal embryos depend on maternally deposited RNA until zygotic genes become transcriptionally active. Before this maternal-to-zygotic transition, many species execute rapid and synchronous cell divisions without growth phases or cell cycle checkpoints. The coordinated onset of transcription, cell cycle lengthening, and cell cycle checkpoints comprise the midblastula transition (MBT). A long-standing model in the frog, Xenopus laevis, posits that MBT timing is controlled by a maternally loaded inhibitory factor that is titrated against the exponentially increasing amount of DNA. To identify MBT regulators, we developed an assay using Xenopus egg extract that recapitulates the activation of transcription only above the DNA-to-cytoplasm ratio found in embryos at the MBT. We used this system to biochemically purify factors responsible for inhibiting transcription below the threshold DNA-to-cytoplasm ratio. This unbiased approach identified histones H3 and H4 as concentration-dependent inhibitory factors. Addition or depletion of H3/H4 from the extract quantitatively shifted the amount of DNA required for transcriptional activation in vitro. Moreover, reduction of H3 protein in embryos induced premature transcriptional activation and cell cycle lengthening, and the addition of H3/H4 shortened post-MBT cell cycles. Our observations support a model for MBT regulation by DNA-based titration and suggest that depletion of free histones regulates the MBT. More broadly, our work shows how a constant concentration DNA binding molecule can effectively measure the amount of cytoplasm per genome to coordinate division, growth, and development.
Keywords: cell size control; early vertebrate development; maternal zygotic transition; systems biology; transcription activation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


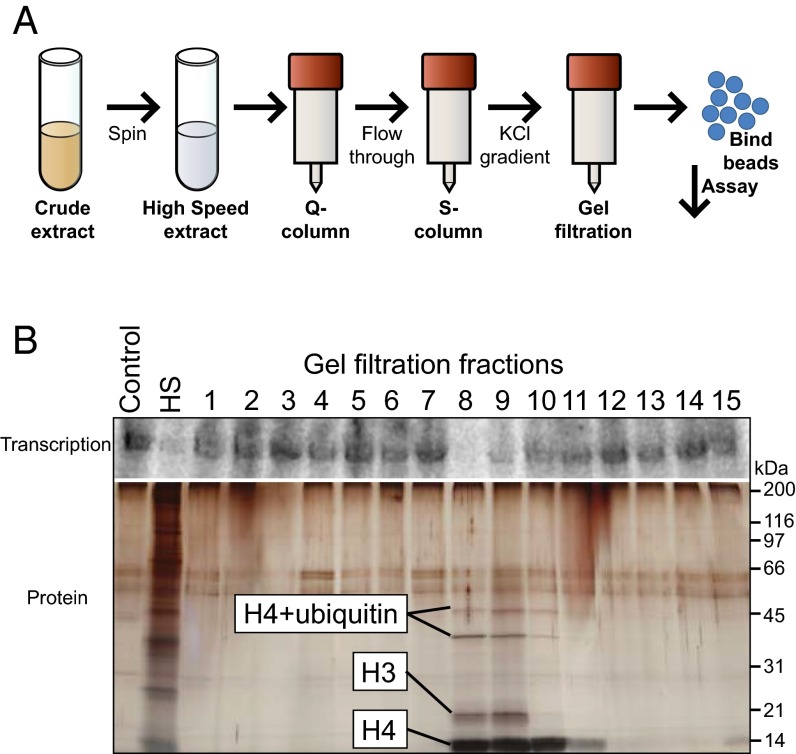



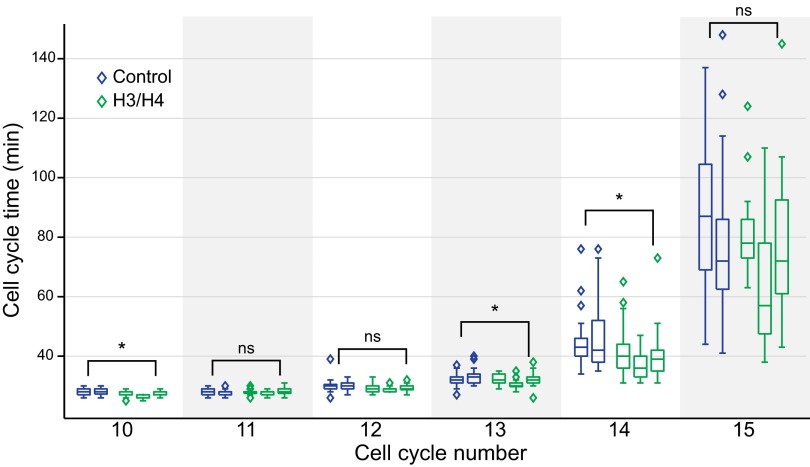

References
-
- Newport J, Kirschner M. A major developmental transition in early Xenopus embryos: II. Control of the onset of transcription. Cell. 1982;30(3):687–696. - PubMed
-
- Newport J, Kirschner M. A major developmental transition in early Xenopus embryos: I. Characterization and timing of cellular changes at the midblastula stage. Cell. 1982;30(3):675–686. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
