

Analysis of 13 cell types reveals evidence for the expression of numerous novel primate- and tissue-specific microRNAs
- PMID: 25713380
- PMCID: PMC4364231
- DOI: 10.1073/pnas.1420955112
Analysis of 13 cell types reveals evidence for the expression of numerous novel primate- and tissue-specific microRNAs
Abstract
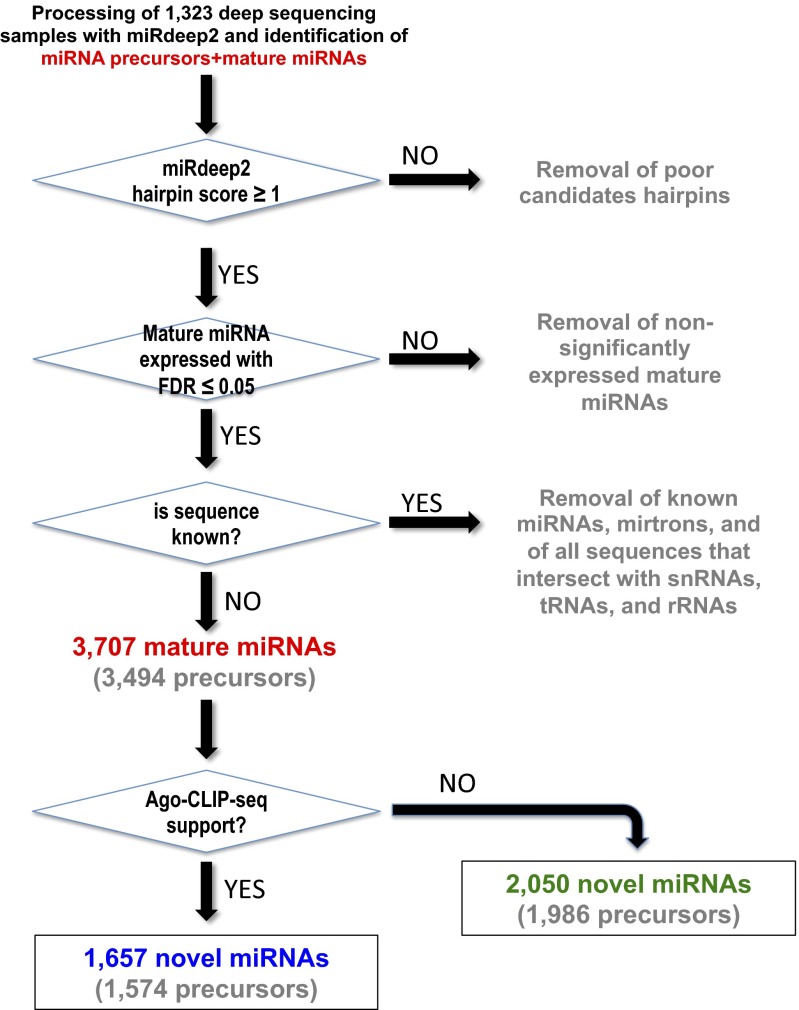
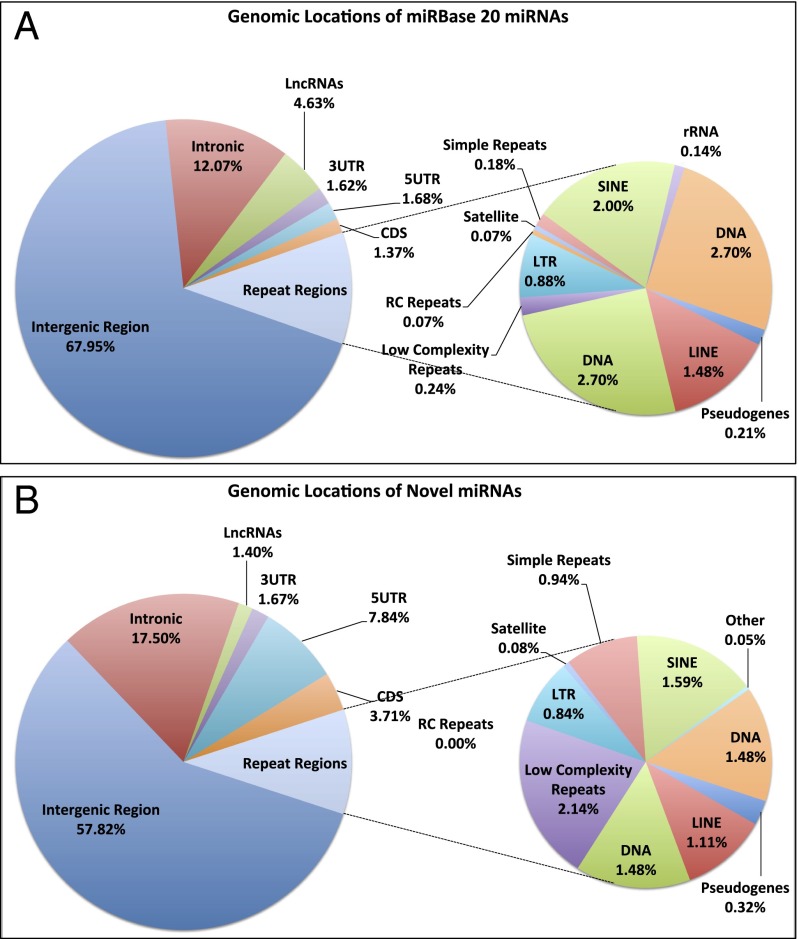
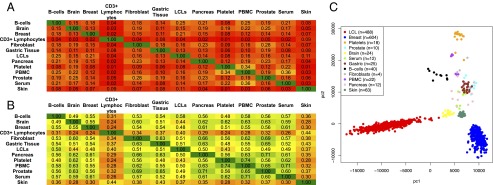
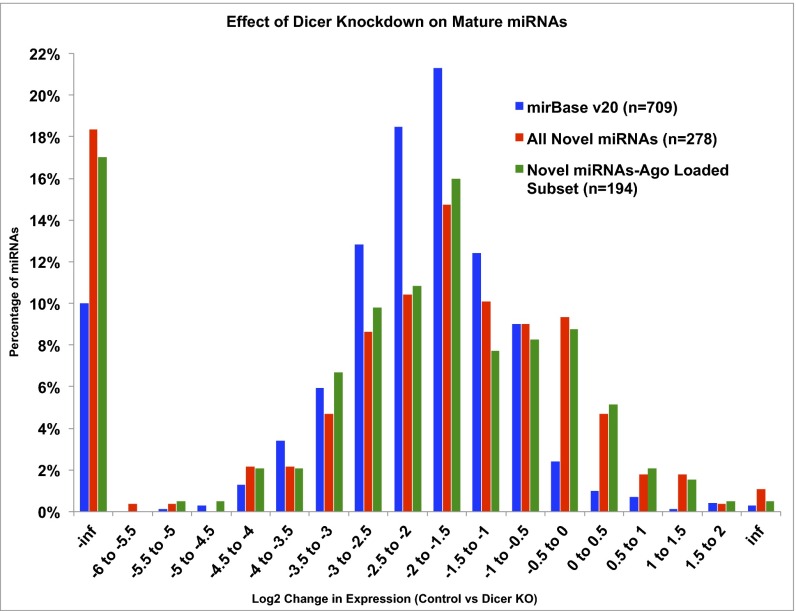
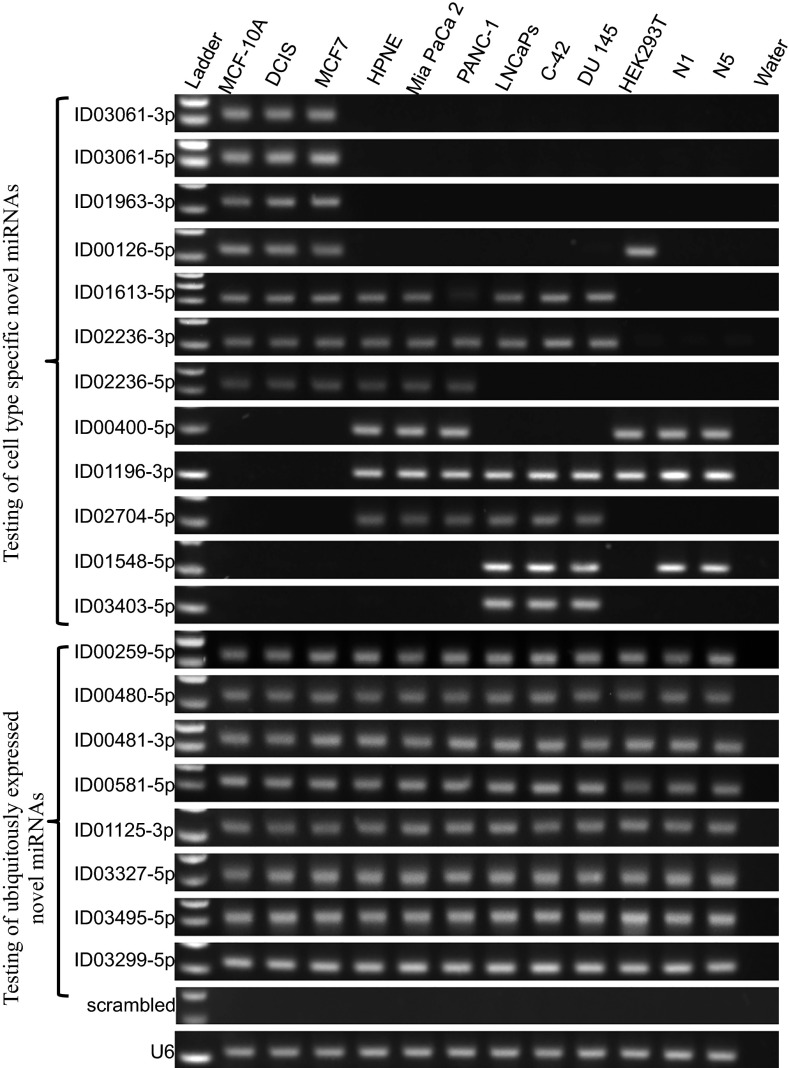
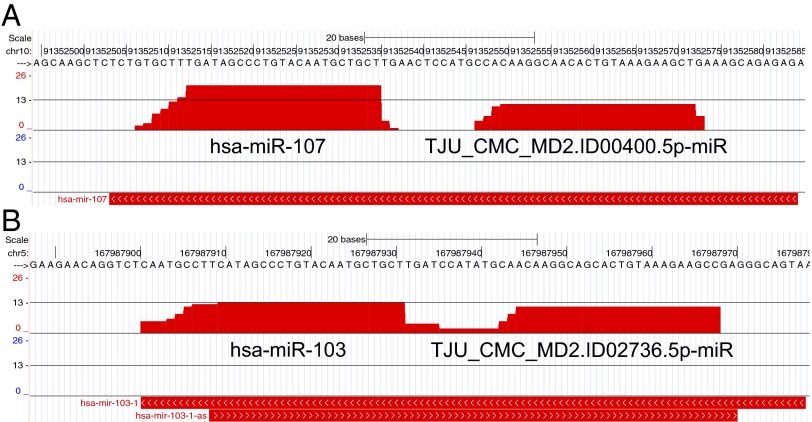
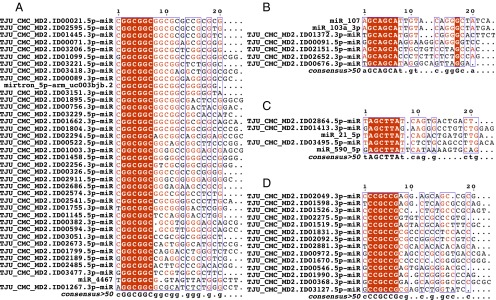
Two decades after the discovery of the first animal microRNA (miRNA), the number of miRNAs in animal genomes remains a vexing question. Here, we report findings from analyzing 1,323 short RNA sequencing samples (RNA-seq) from 13 different human tissue types. Using stringent thresholding criteria, we identified 3,707 statistically significant novel mature miRNAs at a false discovery rate of ≤ 0.05 arising from 3,494 novel precursors; 91.5% of these novel miRNAs were identified independently in 10 or more of the processed samples. Analysis of these novel miRNAs revealed tissue-specific dependencies and a commensurate low Jaccard similarity index in intertissue comparisons. Of these novel miRNAs, 1,657 (45%) were identified in 43 datasets that were generated by cross-linking followed by Argonaute immunoprecipitation and sequencing (Ago CLIP-seq) and represented 3 of the 13 tissues, indicating that these miRNAs are active in the RNA interference pathway. Moreover, experimental investigation through stem-loop PCR of a random collection of newly discovered miRNAs in 12 cell lines representing 5 tissues confirmed their presence and tissue dependence. Among the newly identified miRNAs are many novel miRNA clusters, new members of known miRNA clusters, previously unreported products from uncharacterized arms of miRNA precursors, and previously unrecognized paralogues of functionally important miRNA families (e.g., miR-15/107). Examination of the sequence conservation across vertebrate and invertebrate organisms showed 56.7% of the newly discovered miRNAs to be human-specific whereas the majority (94.4%) are primate lineage-specific. Our findings suggest that the repertoire of human miRNAs is far more extensive than currently represented by public repositories and that there is a significant number of lineage- and/or tissue-specific miRNAs that are uncharacterized.
Keywords: RNA sequencing; isomIRs; microRNAs; noncoding RNA; transcriptome.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Reply to Backes and Keller: Identification of novel tissue-specific and primate-specific human microRNAs.Proc Natl Acad Sci U S A. 2015 Jun 2;112(22):E2851. doi: 10.1073/pnas.1505397112. Epub 2015 May 15. Proc Natl Acad Sci U S A. 2015. PMID: 25979943 Free PMC article. No abstract available.
-
Reanalysis of 3,707 novel human microRNA candidates.Proc Natl Acad Sci U S A. 2015 Jun 2;112(22):E2849-50. doi: 10.1073/pnas.1505017112. Epub 2015 May 15. Proc Natl Acad Sci U S A. 2015. PMID: 25979944 Free PMC article. No abstract available.
References
-
- Bartel DP. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–297. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 GM106047/GM/NIGMS NIH HHS/United States
- P30 CA016672/CA/NCI NIH HHS/United States
- R01 AR 19616/AR/NIAMS NIH HHS/United States
- R01 AG042419/AG/NIA NIH HHS/United States
- 2U19-AI056363-06/20309840/AI/NIAID NIH HHS/United States
- AG028383/AG/NIA NIH HHS/United States
- R01 HL102482/HL/NHLBI NIH HHS/United States
- R01 CA140424/CA/NCI NIH HHS/United States
- R01 MD007880/MD/NIMHD NIH HHS/United States
- P50 CA127297/CA/NCI NIH HHS/United States
- CA140424/CA/NCI NIH HHS/United States
- P30 CA036727/CA/NCI NIH HHS/United States
- R01 HL116713/HL/NHLBI NIH HHS/United States
- NS085830/NS/NINDS NIH HHS/United States
- GM106047/GM/NIGMS NIH HHS/United States
- P30 AI042853/AI/NIAID NIH HHS/United States
- R01 CA099996/CA/NCI NIH HHS/United States
- P30 AG028383/AG/NIA NIH HHS/United States
- R21 NS085830/NS/NINDS NIH HHS/United States
- AG042419/AG/NIA NIH HHS/United States
- P30 CA056036/CA/NCI NIH HHS/United States
- R01 AR019616/AR/NIAMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
