

Regulation of basal promoter activity of the human thiamine pyrophosphate transporter SLC44A4 in human intestinal epithelial cells
- PMID: 25715703
- PMCID: PMC4420793
- DOI: 10.1152/ajpcell.00381.2014
Regulation of basal promoter activity of the human thiamine pyrophosphate transporter SLC44A4 in human intestinal epithelial cells
Erratum in
-
CORRIGENDUM.Am J Physiol Cell Physiol. 2017 Oct 1;313(4):C473. doi: 10.1152/ajpcell.zh0-8195-corr.2017. Am J Physiol Cell Physiol. 2017. PMID: 28993325 Free PMC article. No abstract available.
Abstract
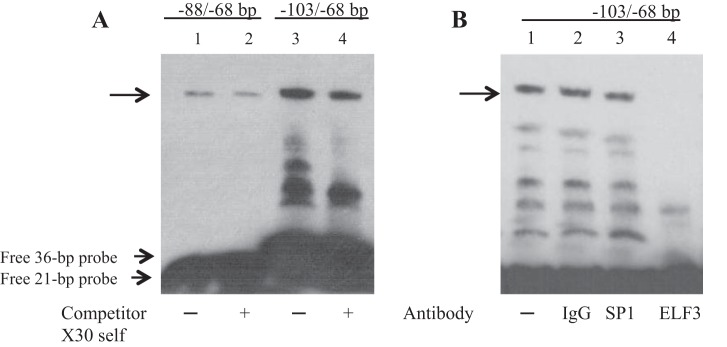
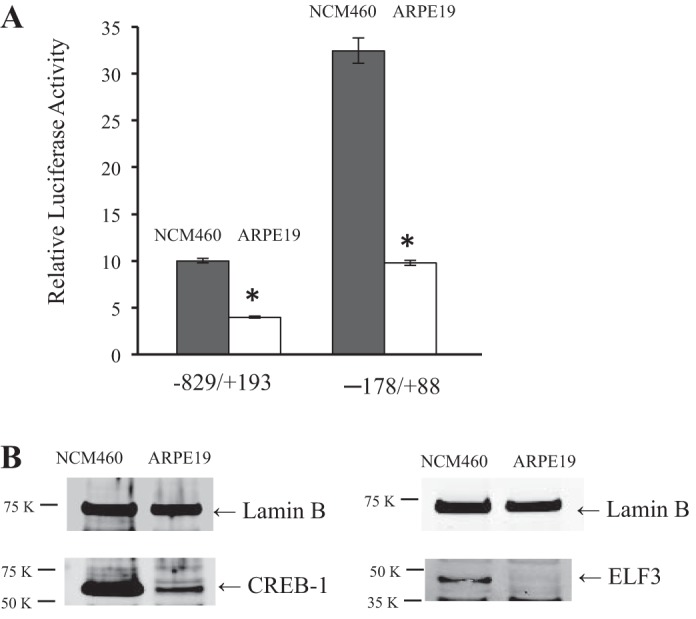
Microbiota of the large intestine synthesize considerable amount of vitamin B1 in the form of thiamine pyrophosphate (TPP). There is a specific high-affinity regulated carrier-mediated uptake system for TPP in human colonocytes (product of the SLC44A4 gene). The mechanisms of regulation of SLC44A4 gene expression are currently unknown. In this study, we characterized the SLC44A4 minimal promoter region and identified transcription factors important for basal promoter activity in colonic epithelial cells. The 5'-regulatory region of the SLC44A4 gene (1,022 bp) was cloned and showed promoter activity upon transient transfection into human colonic epithelial NCM460 cells. With the use of a series of 5'- and 3'-deletion luciferase reporter constructs, the minimal genomic region that required basal transcription of the SLC44A4 gene expression was mapped between nucleotides -178 and +88 (using the distal transcriptional start site as +1). Mutational analysis performed on putative cis-regulatory elements established the involvement of ETS/ELF3 [E26 transformation-specific sequence (ETS) proteins], cAMP-responsive element (CRE), and SP1/GC-box sequence motifs in basal SLC44A4 promoter activity. By means of EMSA, binding of ELF3 and CRE-binding protein-1 (CREB-1) transcription factors to the SLC44A4 minimal promoter was shown. Contribution of CREB into SLC44A4 promoter activity was confirmed using NCM460 cells overexpressing CREB. We also found high expression of ELF3 and CREB-1 in colonic (NCM460) compared with noncolonic (ARPE19) cells, suggesting their possible contribution to colon-specific pattern of SLC44A4 expression. This study represents the first characterization of the SLC44A4 promoter and reports the importance of both ELF3 and CREB-1 transcription factors in the maintenance of basal promoter activity in colonic epithelial cells.
Keywords: SLC44A4; colonocytes; promoter; thiamine pyrophosphate; transcription regulation.
Figures






References
-
- Arumugam M, Raes J, Pelletier E, Le Paslier D, Yamada T, Mende DR, Fernandes GR, Tap J, Bruls T, Batto JM, Bertalan M, Borruel N, Casellas F, Fernandez L, Gautier L, Hansen T, Hattori M, Hayashi T, Kleerebezem M, Kurokawa K, Leclerc M, Levenez F, Manichanh C, Nielsen HB, Nielsen T, Pons N, Poulain J, Qin J, Sicheritz-Ponten T, Tims S, Torrents D, Ugarte E, Zoetendal EG, Wang J, Guarner F, Pedersen O, de Vos WM, Brunak S, Doré J; MetaHIT Consortiuma, Antolín M, Artiguenave F, Blottiere HM, Almeida M, Brechot C, Cara C, Chervaux C, Cultrone A, Delorme C, Denariaz G, Dervyn R, Foerstner KU, Friss C, van de Guchte M, Guedon E, Haimet F, Huber W, van Hylckama-Vlieg J, Jamet A, Juste C, Kaci G, Knol J, Lakhdari O, Layec S, Le Roux K, Maguin E, Mérieux A, Melo Minardi R, M'rini C, Muller J, Oozeer R, Parkhill J, Renault P, Rescigno M, Sanchez N, Sunagawa S, Torrejon A, Turner K, Vandemeulebrouck G, Varela E, Winogradsky Y, Zeller G, Weissenbach J, Ehrlich SD, Bork P. Enterotypes of the human gut microbiome. Nature 473: 174–180, 2011. - PMC - PubMed
-
- Berdanier CD. Advanced Nutrition Micronutrients. Boca Raton, FL: CRC, 1998, p. 80–88.
-
- Bettendorff L, Wins P. Thiamin diphosphate in biological chemistry: new aspects of thiamin metabolism, especially triphosphate derivatives acting other than as cofactors. FEBS J 276: 2917–2925, 2009. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous