

Review
doi: 10.1242/jcs.168021.
Epub 2015 Feb 25.
An active role for basement membrane assembly and modification in tissue sculpting
Affiliations
- PMID: 25717004
- PMCID: PMC4446735
- DOI: 10.1242/jcs.168021
Item in Clipboard
Review
An active role for basement membrane assembly and modification in tissue sculpting
J Cell Sci.
.
Abstract
Basement membranes are a dense, sheet-like form of extracellular matrix (ECM) that underlie epithelia and endothelia, and surround muscle, fat and Schwann cells. Basement membranes separate tissues and protect them from mechanical stress. Although traditionally thought of as a static support structure, a growing body of evidence suggests that dynamic basement membrane deposition and modification instructs coordinated cellular behaviors and acts mechanically to sculpt tissues. In this Commentary, we highlight recent studies that support the idea that far from being a passive matrix, basement membranes play formative roles in shaping tissues.
© 2015. Published by The Company of Biologists Ltd.
Figures
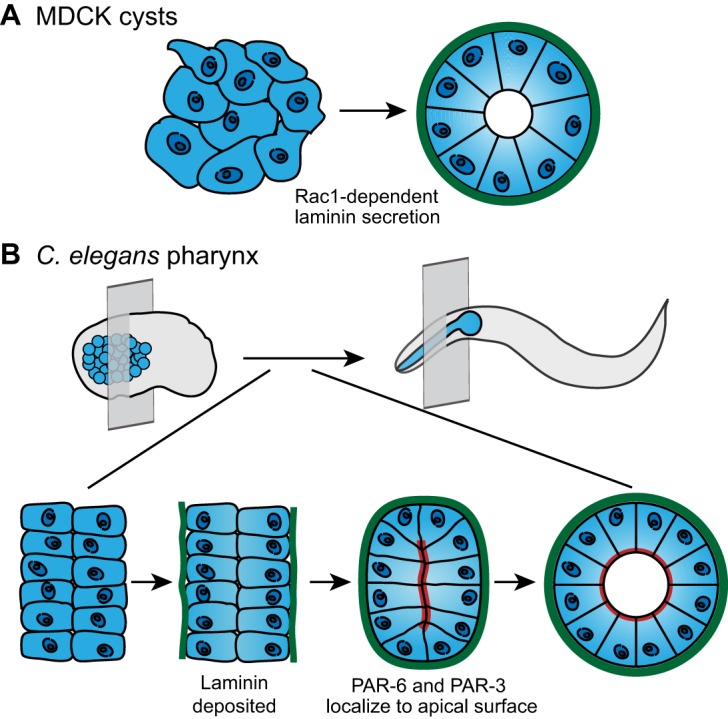
Basement membrane deposition instructs cell polarization and tissue shape. (A) MDCK cells (left) secrete laminin (green) in a Rac1-dependent manner. Laminin accumulates on the basal surface of the cells and directs coordinated apical-basal polarization, which results in formation of a cyst with an apical lumen (right). (B) In the C. elegans embryo (top left), the pharyngeal precursors (blue) organize into two plates of cells (bottom left; perspective indicated by transverse plane in top left). Laminin (green) accumulates on the basal surface of the pharyngeal precursors (middle left). Laminin assembly on the basal cell surface precedes the apical localization of polarity proteins PAR-6 and PAR-3 (red; middle right), as well as apical lumen formation (bottom right). Lumen formation is necessary to shape a functional pharynx in the larva and adult C. elegans (top right). Data are from O'Brien et al. and Rasmussen et al. (O'Brien et al., 2001; Rasmussen et al., 2012) and have been adapted with permission.
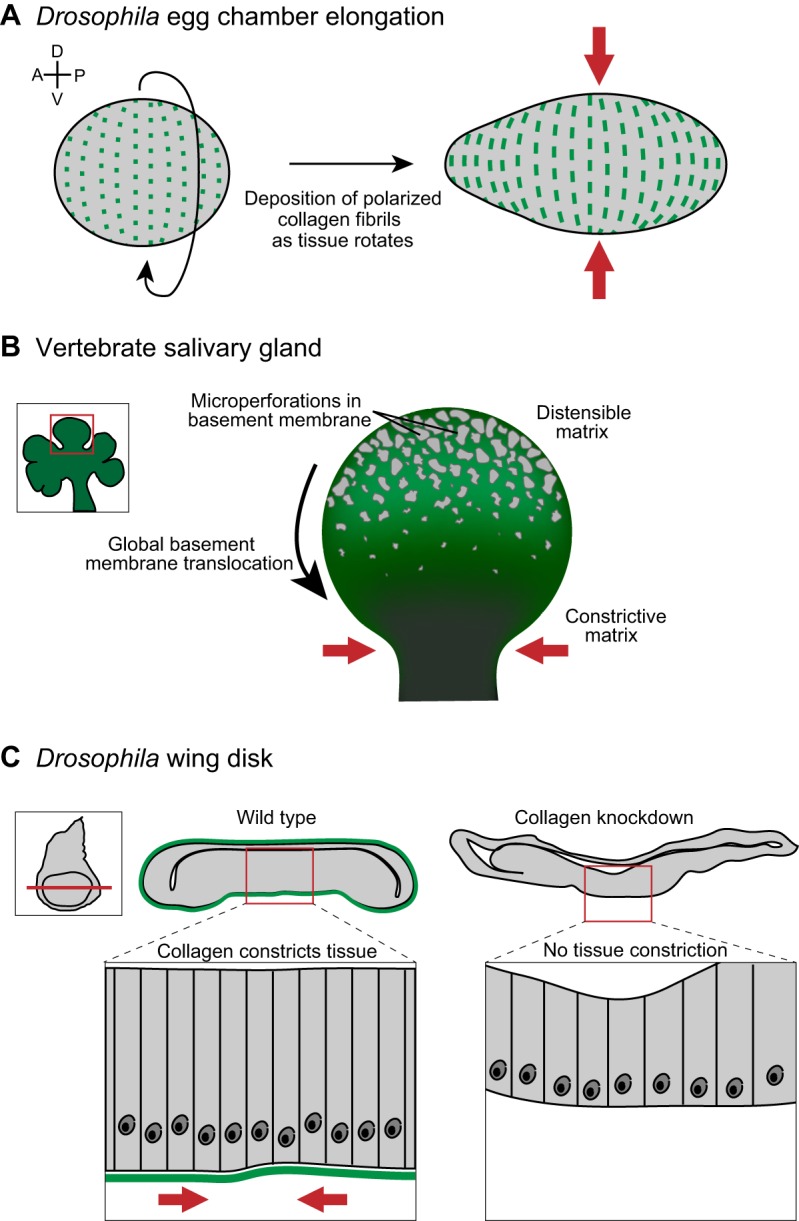
Collagen within basement membranes constricts and contours tissues. (A) Prior to tissue elongation, the Drosophila egg chamber initiates global tissue revolutions, directing the deposition of oriented collagen fibrils (green) as it rotates (left). The collagen fibrils constrict egg chamber growth in the dorsal-ventral (D-V) tissue axis (red arrows), forcing elongation along the anterior-posterior (A-P) axis (right). (B) Inset shows a developing salivary gland, with a single bud (red box in inset) highlighted on the right. Imaging of basement membrane (green) dynamics during salivary gland branching morphogenesis revealed a perforated, distensible matrix at the bud tip, which facilitates tissue growth. During bud tip growth, the basement membrane translocates from the bud tip to the stalk, where basement membrane movement slows and collagen accumulates. This results in a constrictive basement membrane corset around the stalk (red arrows), which stabilizes and likely restricts growth at the stalk. (C) A tissue-specific RNAi strategy revealed that collagen is required to shape the imaginal wing disc in Drosophila (inset). Cross sections (position indicated by red line in inset) through a wild-type (left) and collagen-knockdown (right) wing disc show that the wild-type wing disc is more compact than the collagen-knockdown wing disc. A magnified view (below) illustrates the highly ordered columnar epithelium in wild type compared to a flattened epithelium following collagen knockdown. Data are from Haigo and Bilder, 2011, Harunaga et al. and Pastor-Pareja and Xu (Haigo and Bilder, 2011; Harunaga et al., 2014; Pastor-Pareja and Xu, 2011) and have been adapted with permission.
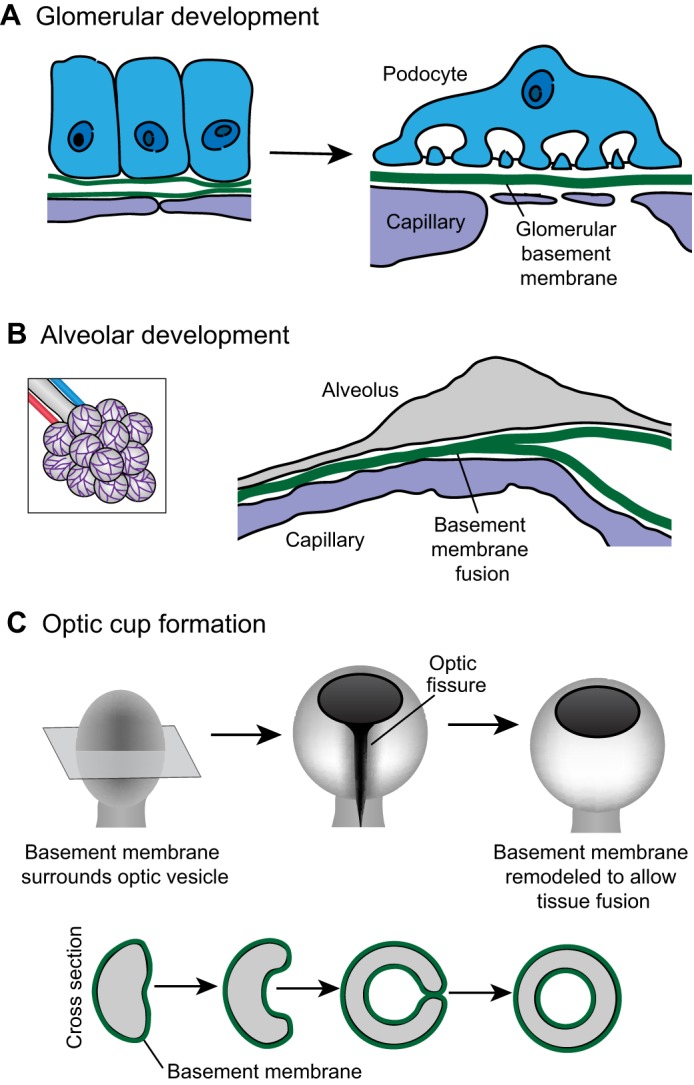
Basement-membrane–basement-membrane interactions. (A) As the kidney develops, the epithelial podocyte (blue) and the vascular endothelium (purple) each synthesize basement membrane (left). These individual basement membranes fuse to form the glomerular basement membrane (right). (B) During development of the alveolus (left; a magnified view is shown on the right), the capillary (purple) basement membrane and the alveolar (gray) basement membrane fuse to form a single sheet of basement membrane (C) The optic cup initiates as an epithelial bud called the optic vesicle (left). Shown in cross section (bottom), the center of the optic vesicle invaginates and the sides of the optic vesicle grow circumferentially until they meet to close the optic fissure and form a continuous circular cup (right). When the basement membrane-encased sides of the optic cup meet (middle right), the basement membrane is remodeled to allow direct cell–cell contract (right). Data are from Vaccaro and Brody (©1981 Vaccaro and Brody. Journal of Cell Biology. 91:427–437. doi: 10.1083/jcb.91.2.427) and St John and Abrahamson (St John and Abrahamson, 2001) and have been adapted with permission.
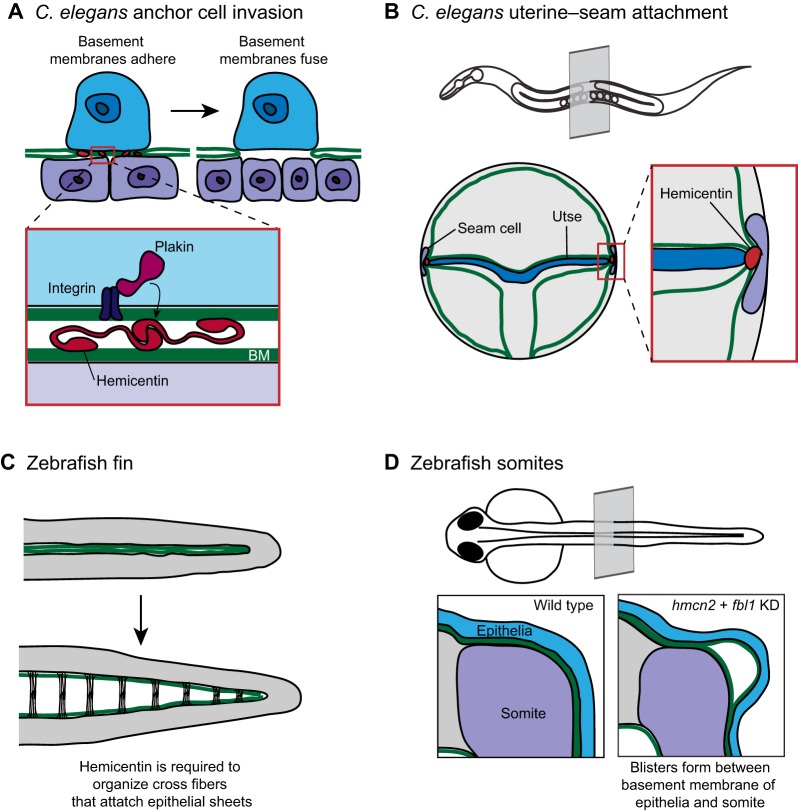
Hemicentin regulates basement membrane-basement membrane adhesion. (A) The C. elegans uterine anchor cell (blue) breaches the juxtaposed gonadal and ventral basement membranes (green) and contacts the underlying vulval epithelium (purple). Just prior to basement membrane breach (left), the individual basement membranes are connected by a B-LINK adhesion structure with its molecular regulators shown in the inset. Extracellular hemicentin (red) localizes between the basement membranes. Intracellular plakin (magenta) and integrin (dark purple) are required to form a functional B-LINK and might stabilize the adhesion by connecting the B-LINK to the anchor cell cytoskeleton. (B) The hemicentin, plakin and integrin-dependent B-LINK is also required to attach the C. elegans seam cell (purple) to the uterine utse cell (blue). A transverse plane through an adult worm (top) indicates the location of the uterine–seam attachment. (C) The zebrafish fin originates as two abutting sheets of epithelia (top). During fin development, these sheets separate but remain attached through extracellular cross-fibers (bottom). Hemicentin is required to organize the cross fibers that attach the epithelial sheets. (D) A cross section through a zebrafish larva shows the juxtaposed basement membranes surrounding the somite (purple) and the epithelia (blue). Following knockdown of the hemicentin ortholog hmcn2 and the fibulin family member fbl1, blisters develop between the somite basement membrane and the epithelial basement membrane (right). Data from Morrissey et al. and Feitosa et al. (Morrissey et al., 2014; Feitosa et al., 2012) and have been adapted with permission.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
