

Functional characterization of IgA-targeted bacterial taxa from undernourished Malawian children that produce diet-dependent enteropathy
- PMID: 25717097
- PMCID: PMC4423598
- DOI: 10.1126/scitranslmed.aaa4877
Functional characterization of IgA-targeted bacterial taxa from undernourished Malawian children that produce diet-dependent enteropathy
Abstract
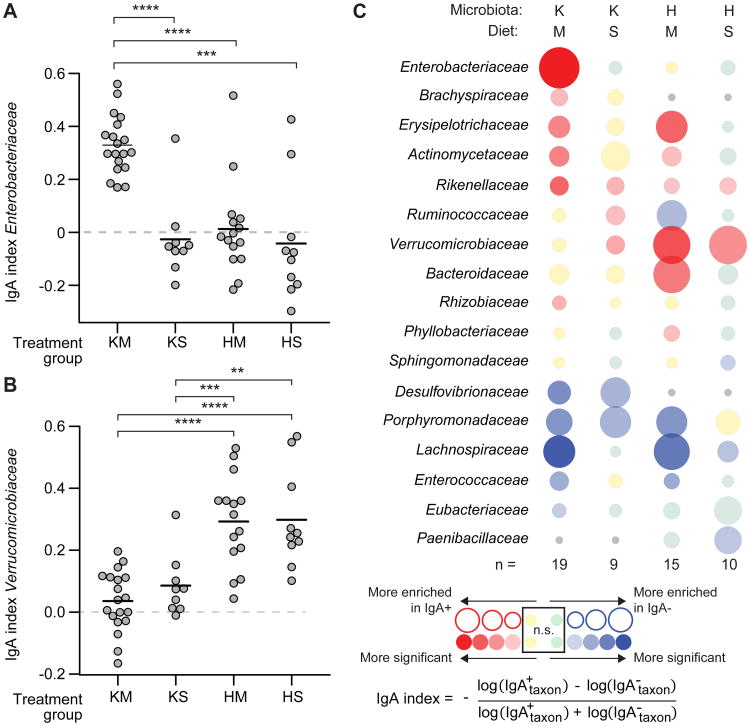
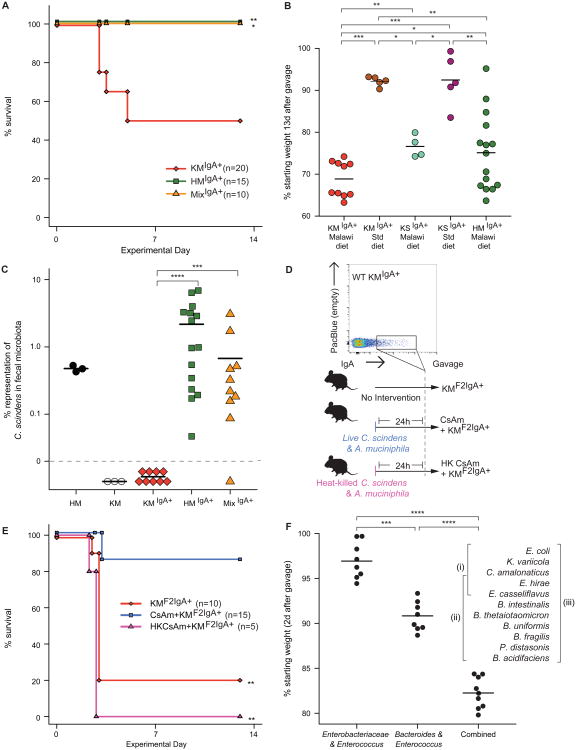
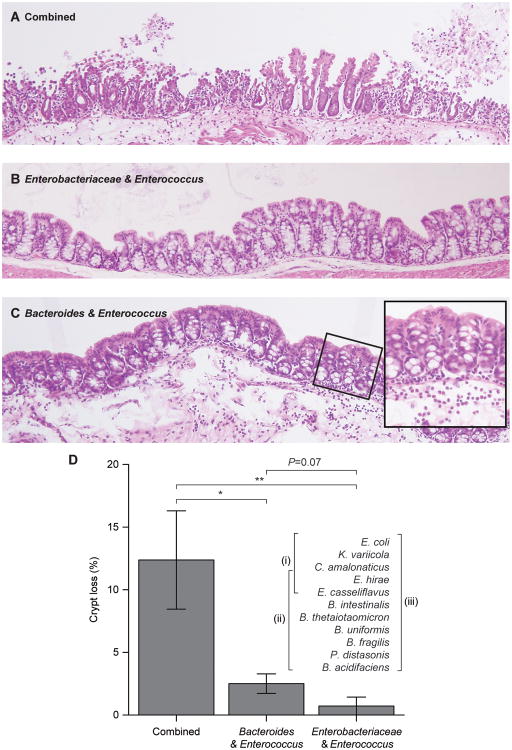
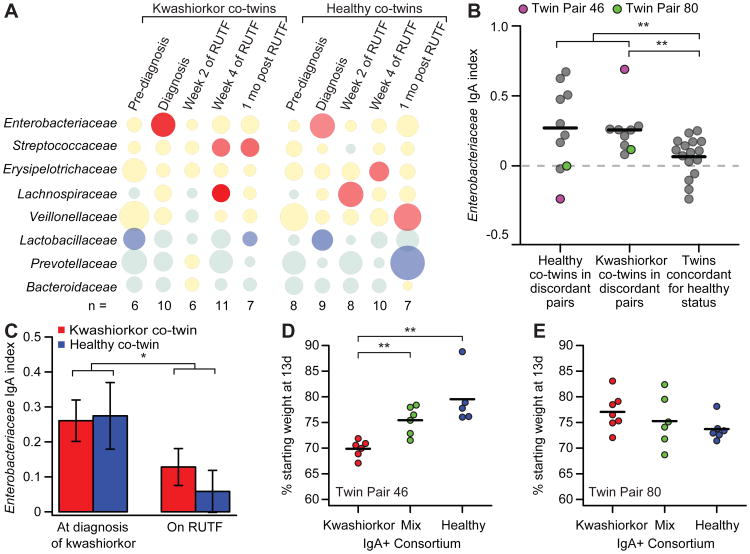
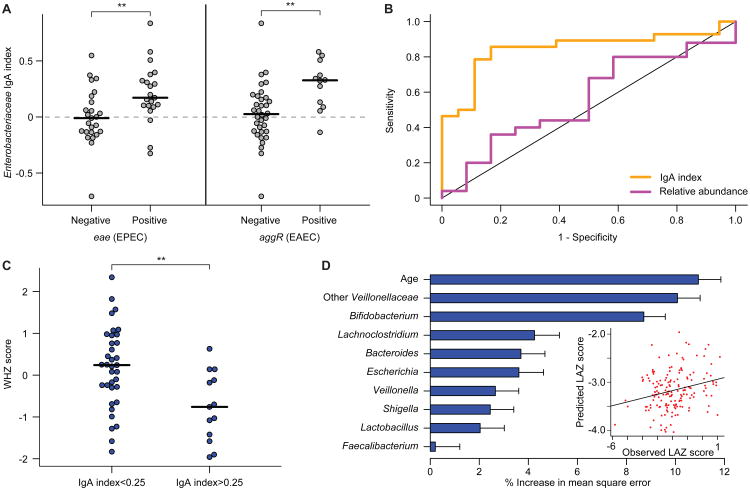
To gain insights into the interrelationships among childhood undernutrition, the gut microbiota, and gut mucosal immune/barrier function, we purified bacterial strains targeted by immunoglobulin A (IgA) from the fecal microbiota of two cohorts of Malawian infants and children. IgA responses to several bacterial taxa, including Enterobacteriaceae, correlated with anthropometric measurements of nutritional status in longitudinal studies. The relationship between IgA responses and growth was further explained by enteropathogen burden. Gnotobiotic mouse recipients of an IgA(+) bacterial consortium purified from the gut microbiota of undernourished children exhibited a diet-dependent enteropathy characterized by rapid disruption of the small intestinal and colonic epithelial barrier, weight loss, and sepsis that could be prevented by administering two IgA-targeted bacterial species from a healthy microbiota. Dissection of a culture collection of 11 IgA-targeted strains from an undernourished donor, sufficient to transmit these phenotypes, disclosed that Enterobacteriaceae interacted with other consortium members to produce enteropathy. These findings indicate that bacterial targets of IgA responses have etiologic, diagnostic, and therapeutic implications for childhood undernutrition.
Copyright © 2015, American Association for the Advancement of Science.
Figures
References
-
- Levels and Trends in Child Malnutrition, UNICEF-WHO-The World Bank Joint Child Malnutrition Estimates. 2012 URL: http://www.who.int/nutgrowthdb/estimates/en/
-
- Black RE, Victora CG, Walker SP, Bhutta ZA, Christian P, de Onis M, Ezzati M, Grantham-McGregor S, Katz J, Martorell R, Uauy R. Maternal and child undernutrition and overweight in low-income and middle-income countries. Lancet. 2013;382:427–451. - PubMed
-
- Ruel MT, Alderman H. Nutrition-sensitive interventions and programmes: how can they help to accelerate progress in improving maternal and child nutrition? Lancet. 2013;382:536–551. - PubMed
-
- Yatsunenko T, Rey FE, Manary MJ, Trehan I, Dominguez-Bello MG, Contreras M, Magris M, Hidalgo G, Baldassano RN, Anokhin AP, Heath AC, Warner B, Reeder J, Kuczynski J, Caporaso JG, Lozupone CA, Lauber C, Clemente JC, Knights D, Knight R, Gordon JI. Human gut microbiome viewedacross age and geography. Nature. 2012;486:222–227. - PMC - PubMed
-
- Subramanian S, Huq S, Yatsunenko T, Haque R, Mahfuz M, Alam MA, Benezra A, DeStefano J, Meier MF, Muegge BD, Barratt MJ, VanArendonk LG, Zhang Q, Province MA, Petri WA, Jr, Ahmed T, Gordon JI. Persistent gut microbiota immaturity in malnourished Bangladeshi children. Nature. 2014;510:417–421. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
