

Unraveling the mechanistic features of RNA polymerase II termination by the 5'-3' exoribonuclease Rat1
- PMID: 25722373
- PMCID: PMC4357727
- DOI: 10.1093/nar/gkv133
Unraveling the mechanistic features of RNA polymerase II termination by the 5'-3' exoribonuclease Rat1
Abstract
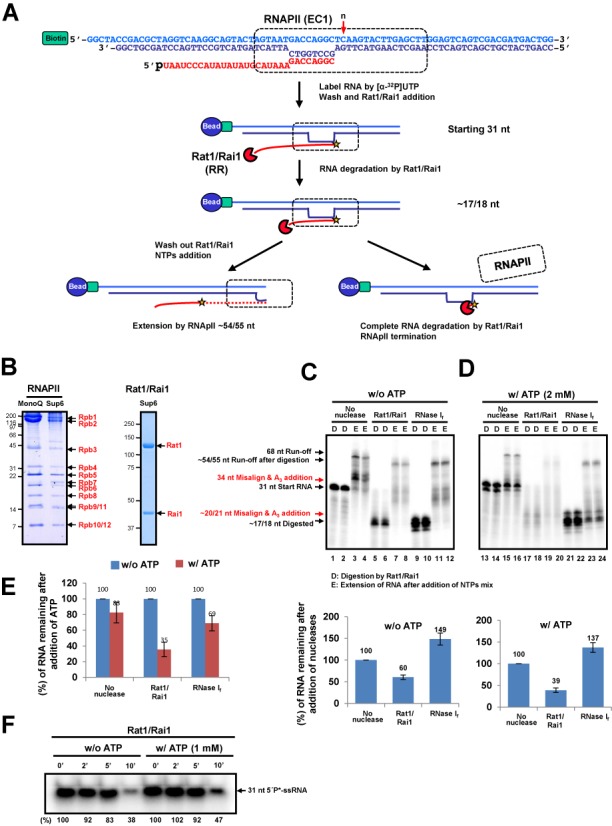
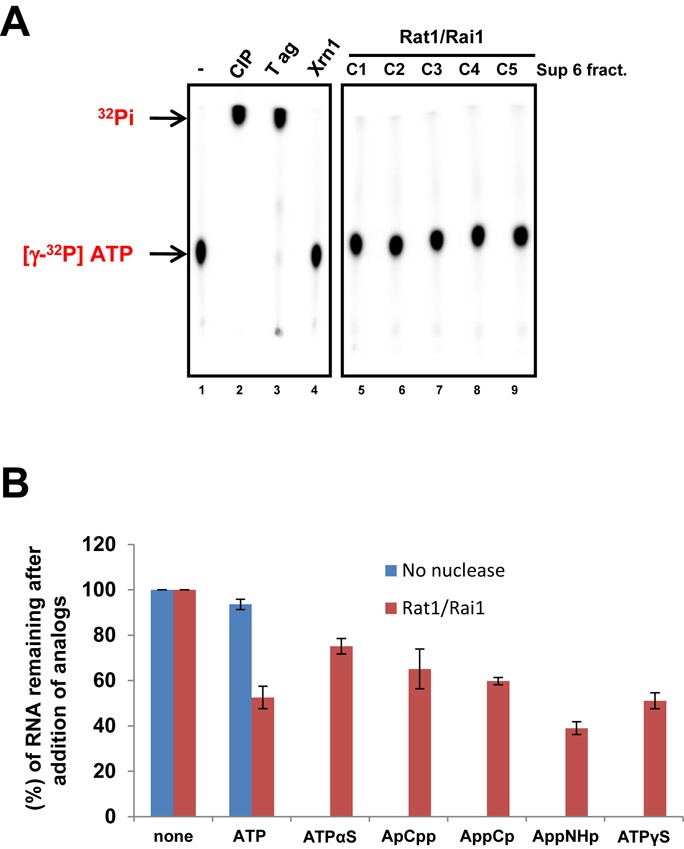
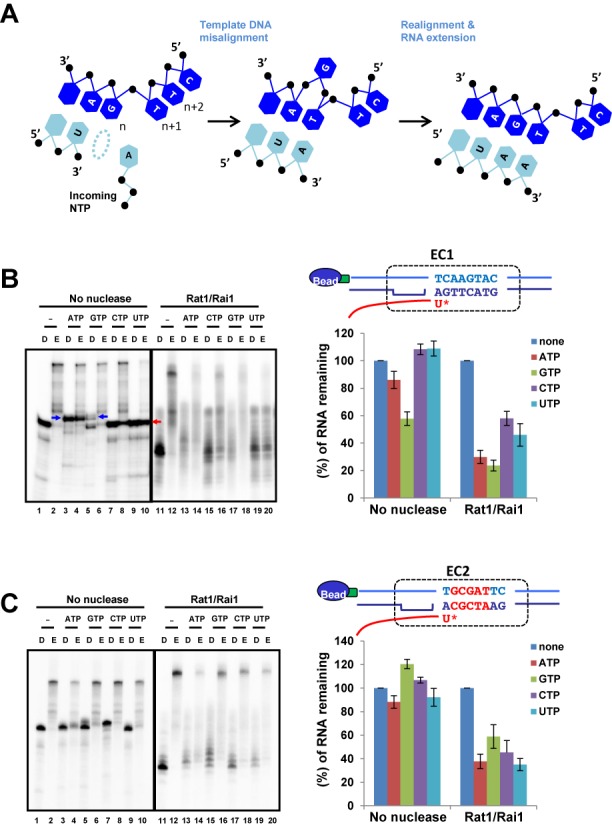
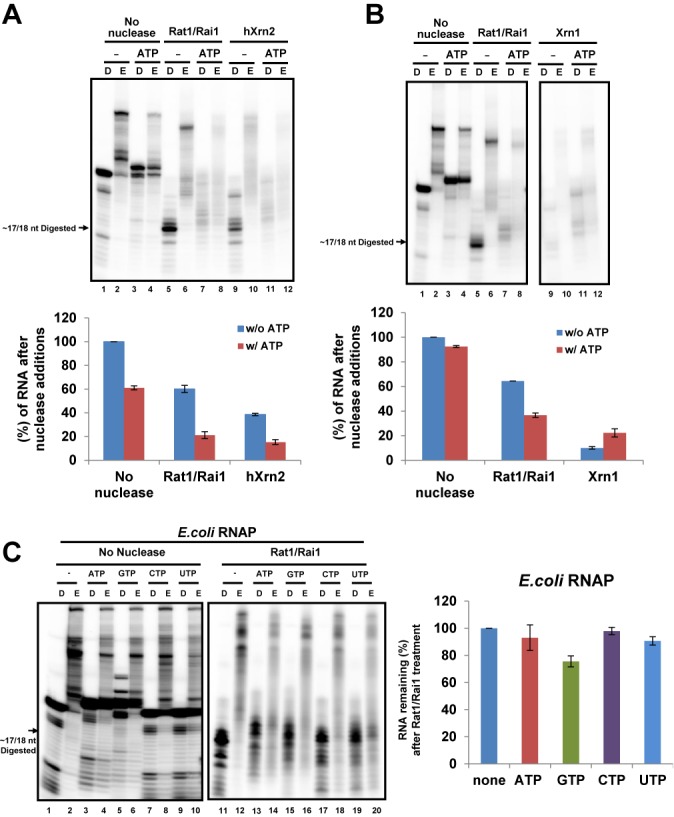
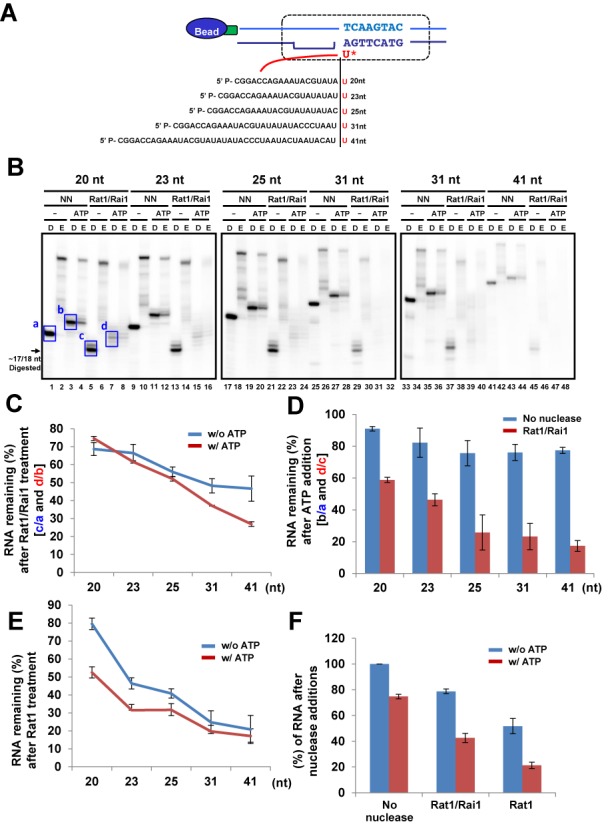
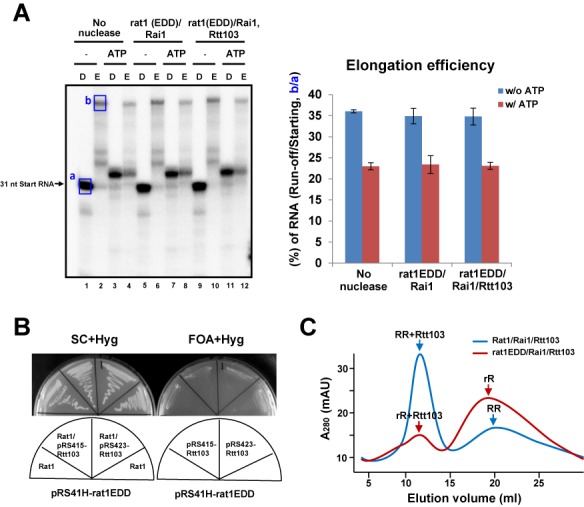
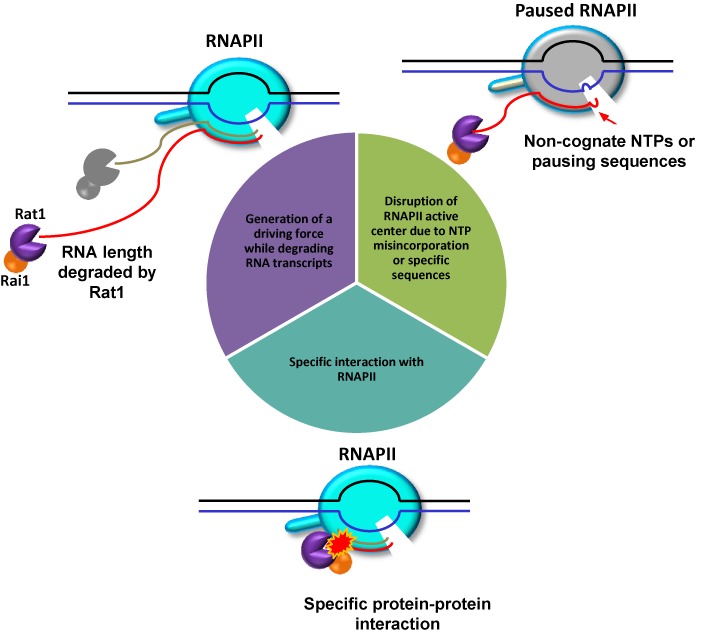
Within a complex with Rai1, the 5'-3' exoribonuclease Rat1 promotes termination of RNA polymerase II (RNAPII) on protein-coding genes, but its underlying molecular mechanism is still poorly understood. Using in vitro transcription termination assays, we have found that RNAPII is prone to more effective termination by Rat1/Rai1 when its catalytic site is disrupted due to NTP misincorporation, implying that paused RNAPII, which is often found in vivo near termination sites, could adopt a similar configuration to Rat1/Rai1 and trigger termination. Intriguingly, yeast Rat1/Rai1 does not terminate Escherichia coli RNAP, implying that a specific interaction between Rat1/Rai1 and RNAPII may be required for termination. Furthermore, the efficiency of termination increases as the RNA transcript undergoing degradation by Rat1 gets longer, which suggests that Rat1 may generate a driving force for dissociating RNAPII from the template while degrading the nascent transcripts to catch up to the polymerase. These results indicate that multiple mechanistic features contribute to Rat1-mediated termination of RNAPII.
© The Author(s) 2015. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Kim M., Vasiljeva L., Rando O.J., Zhelkovsky A., Moore C., Buratowski S. Distinct pathways for snoRNA and mRNA termination. Mol. Cell. 2006;24:723–734. - PubMed
-
- Steinmetz E.J., Conrad N.K., Brow D.A., Corden J.L. RNA-binding protein Nrd1 directs poly(A)-independent 3′-end formation of RNA polymerase II transcripts. Nature. 2001;413:327–331. - PubMed
-
- Steinmetz E.J., Warren C.L., Kuehner J.N., Panbehi B., Ansari A.Z., Brow D.A. Genome-wide distribution of yeast RNA polymerase II and its control by Sen1 helicase. Mol. Cell. 2006;24:735–746. - PubMed
-
- Kim M., Krogan N.J., Vasiljeva L., Rando O.J., Nedea E., Greenblatt J.F., Buratowski S. The yeast Rat1 exonuclease promotes transcription termination by RNA polymerase II. Nature. 2004;432:517–522. - PubMed
-
- West S., Gromak N., Proudfoot N.J. Human 5′ –> 3′ exonuclease Xrn2 promotes transcription termination at co-transcriptional cleavage sites. Nature. 2004;432:522–525. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
