

Mechanism of human antibody-mediated neutralization of Marburg virus
- PMID: 25723164
- PMCID: PMC4344968
- DOI: 10.1016/j.cell.2015.01.031
Mechanism of human antibody-mediated neutralization of Marburg virus
Abstract
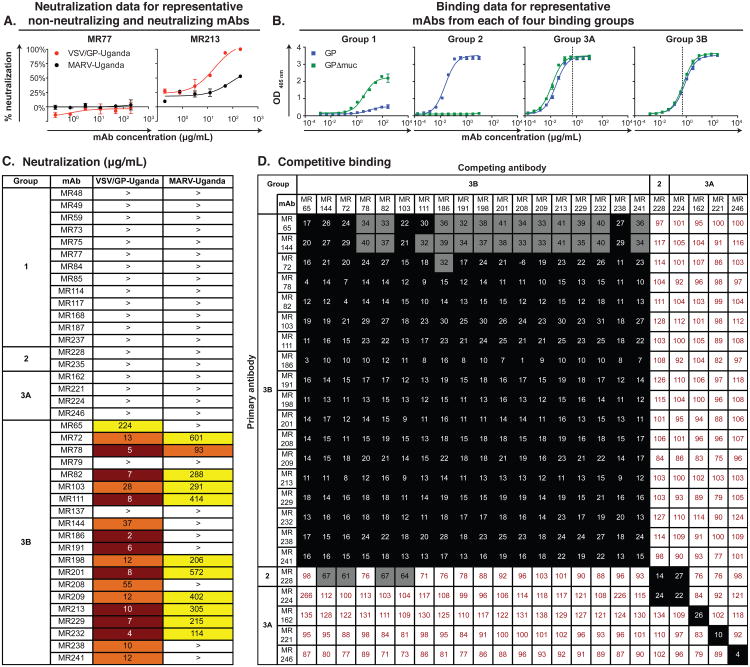
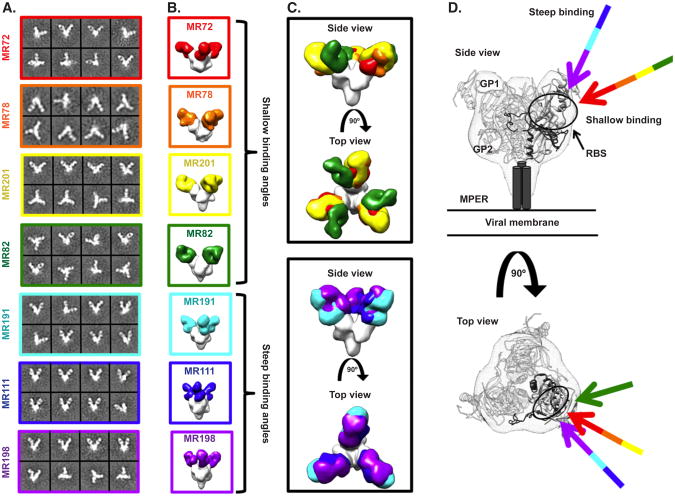
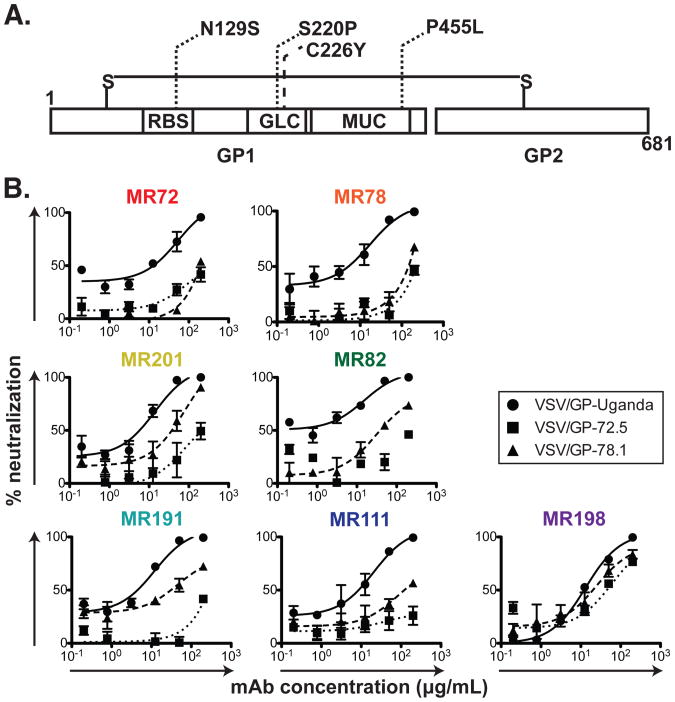
The mechanisms by which neutralizing antibodies inhibit Marburg virus (MARV) are not known. We isolated a panel of neutralizing antibodies from a human MARV survivor that bind to MARV glycoprotein (GP) and compete for binding to a single major antigenic site. Remarkably, several of the antibodies also bind to Ebola virus (EBOV) GP. Single-particle EM structures of antibody-GP complexes reveal that all of the neutralizing antibodies bind to MARV GP at or near the predicted region of the receptor-binding site. The presence of the glycan cap or mucin-like domain blocks binding of neutralizing antibodies to EBOV GP, but not to MARV GP. The data suggest that MARV-neutralizing antibodies inhibit virus by binding to infectious virions at the exposed MARV receptor-binding site, revealing a mechanism of filovirus inhibition.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
-
- Carragher B, Kisseberth N, Kriegman D, Milligan RA, Potter CS, Pulokas J, Reilein A. Leginon: An automated system for acquisition of images from vitreous ice specimens. J Struct Biol. 2000;132:33–45. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- UL1 TR000445/TR/NCATS NIH HHS/United States
- UL1 TR000445-06/TR/NCATS NIH HHS/United States
- DK058404/DK/NIDDK NIH HHS/United States
- P41 GM103310/GM/NIGMS NIH HHS/United States
- U19 AI109711/AI/NIAID NIH HHS/United States
- P30 DK058404/DK/NIDDK NIH HHS/United States
- P30 CA68485/CA/NCI NIH HHS/United States
- 1U19AI109711/AI/NIAID NIH HHS/United States
- R01 AI089498/AI/NIAID NIH HHS/United States
- U19 AI109762/AI/NIAID NIH HHS/United States
- T32 AI007244/AI/NIAID NIH HHS/United States
- UL1 RR024975-01/RR/NCRR NIH HHS/United States
- T32 HL069765/HL/NHLBI NIH HHS/United States
- P30 CA068485/CA/NCI NIH HHS/United States
- R01AI089498/AI/NIAID NIH HHS/United States
- GM103310/GM/NIGMS NIH HHS/United States
- U19AI109762/AI/NIAID NIH HHS/United States
- UL1 RR024975/RR/NCRR NIH HHS/United States
- U01 AI082156/AI/NIAID NIH HHS/United States
- U01AI082156/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
