

Natural grouping of neural responses reveals spatially segregated clusters in prearcuate cortex
- PMID: 25728571
- PMCID: PMC4366683
- DOI: 10.1016/j.neuron.2015.02.014
Natural grouping of neural responses reveals spatially segregated clusters in prearcuate cortex
Abstract
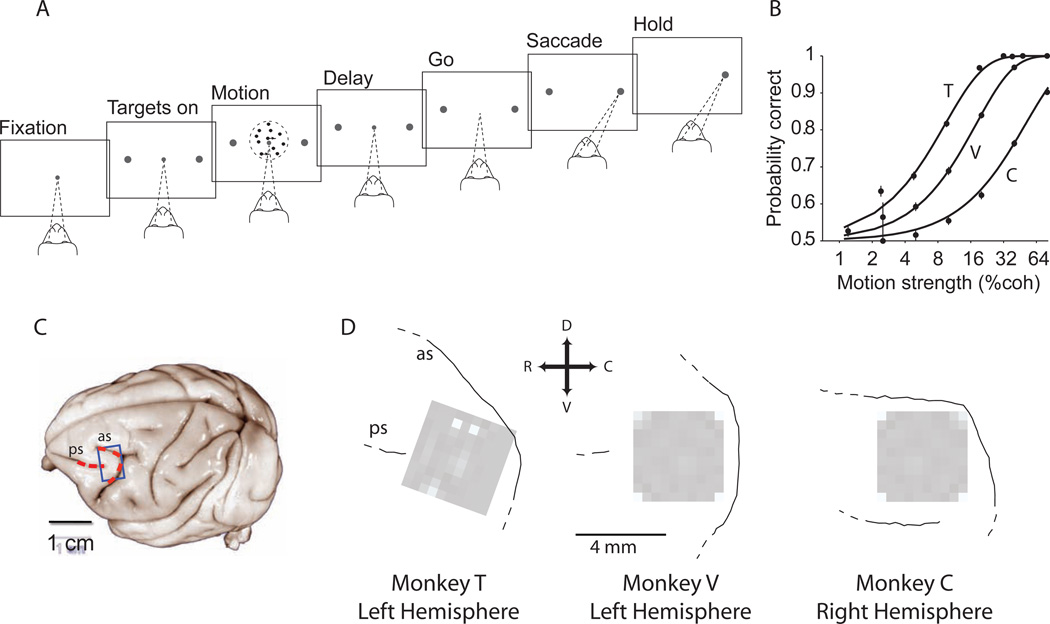
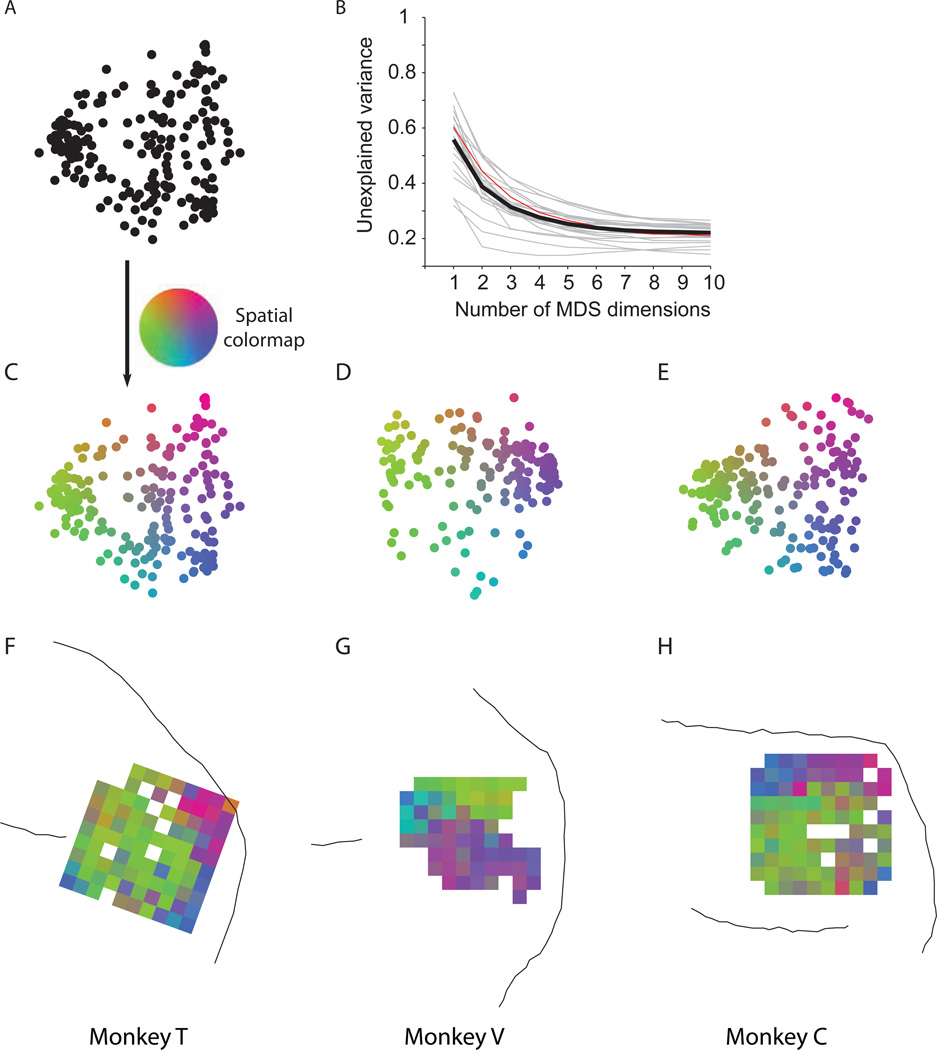
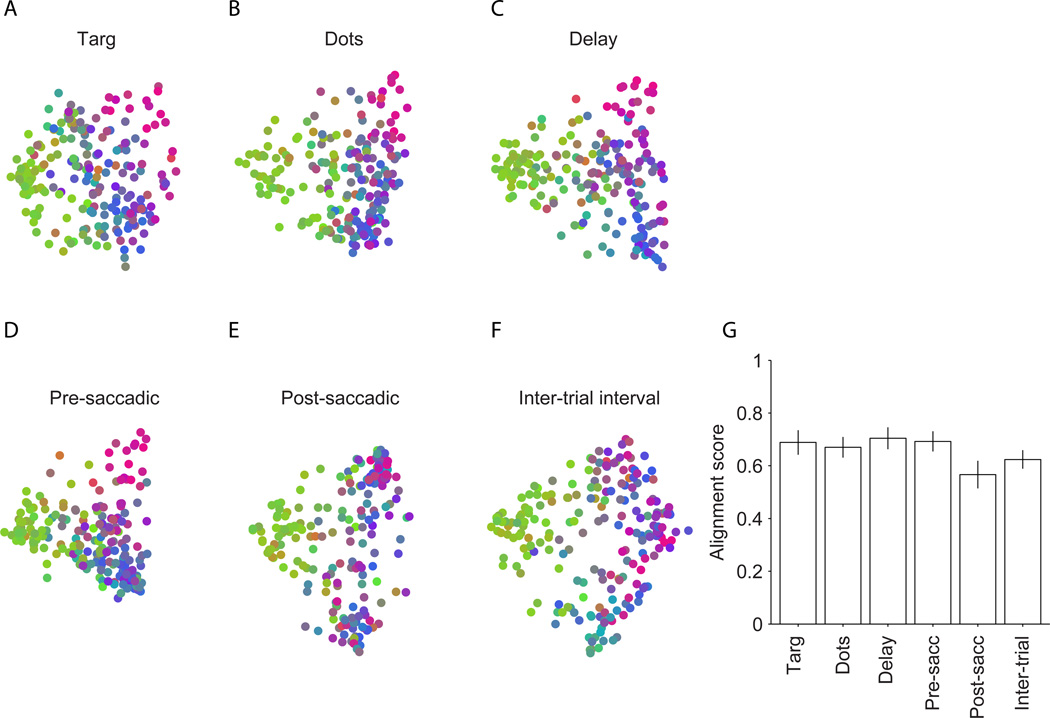
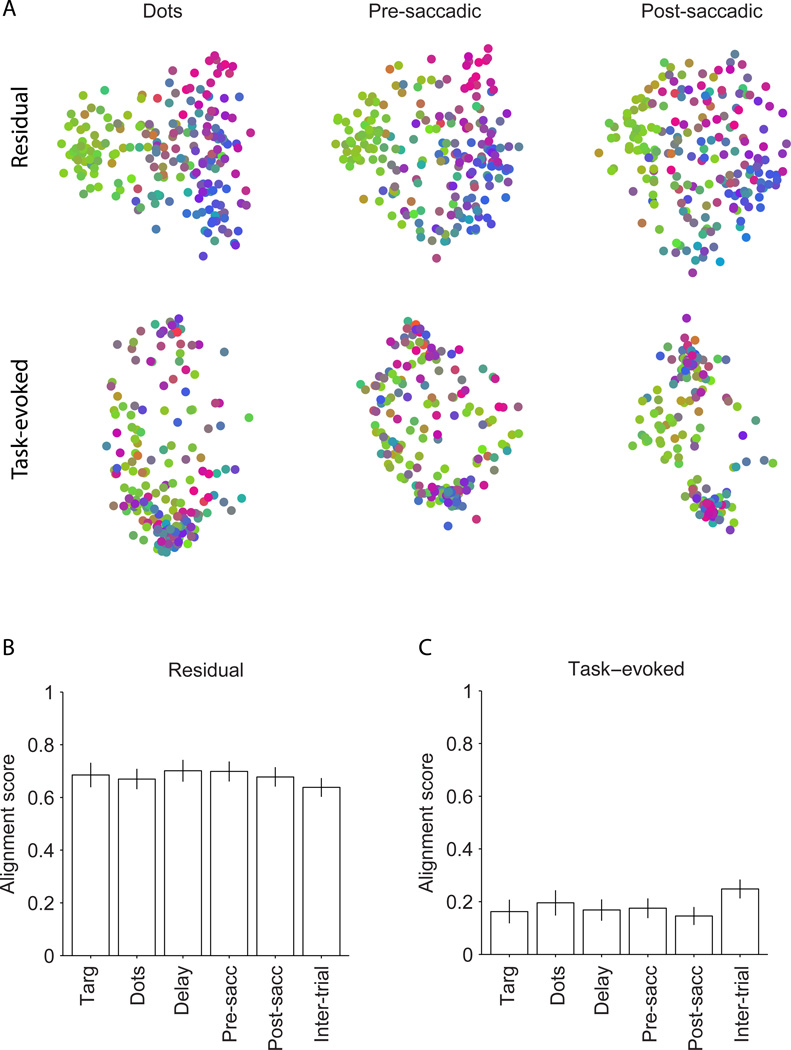
A fundamental challenge in studying the frontal lobe is to parcellate this cortex into "natural" functional modules despite the absence of topographic maps, which are so helpful in primary sensory areas. Here we show that unsupervised clustering algorithms, applied to 96-channel array recordings from prearcuate gyrus, reveal spatially segregated subnetworks that remain stable across behavioral contexts. Looking for natural groupings of neurons based on response similarities, we discovered that the recorded area includes at least two spatially segregated subnetworks that differentially represent behavioral choice and reaction time. Importantly, these subnetworks are detectable during different behavioral states and, surprisingly, are defined better by "common noise" than task-evoked responses. Our parcellation process works well on "spontaneous" neural activity, and thus bears strong resemblance to the identification of "resting-state" networks in fMRI data sets. Our results demonstrate a powerful new tool for identifying cortical subnetworks by objective classification of simultaneously recorded electrophysiological activity.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
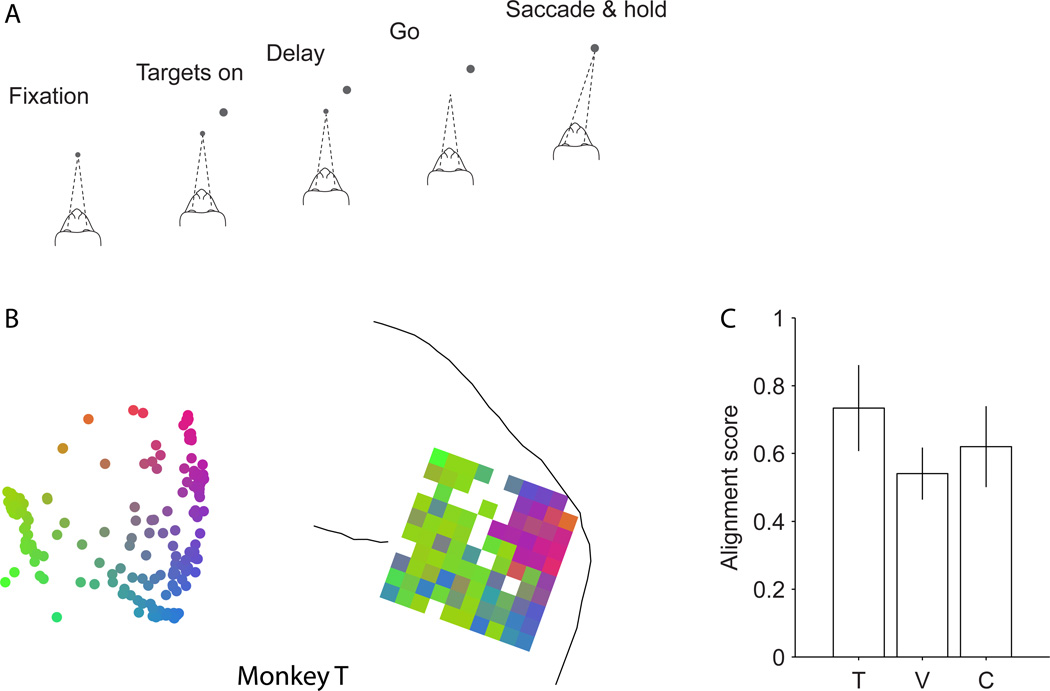
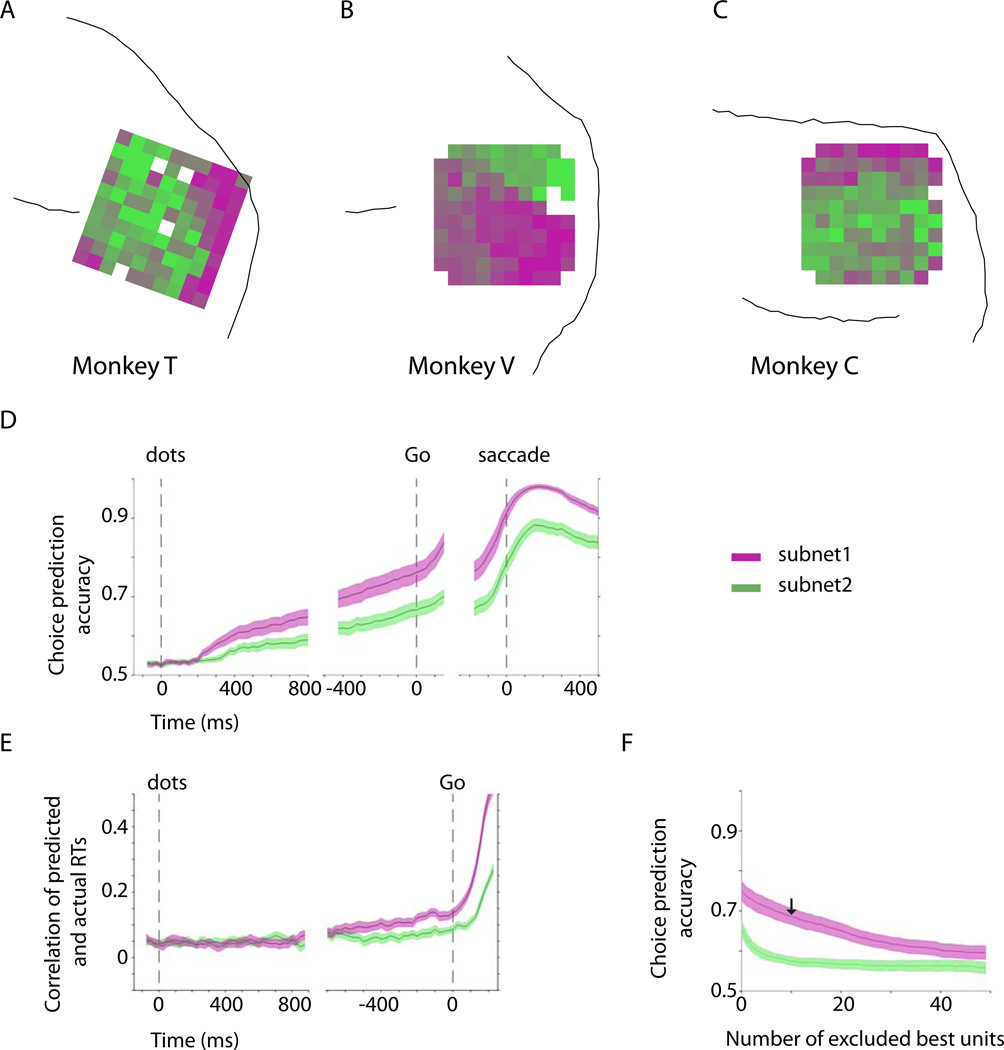
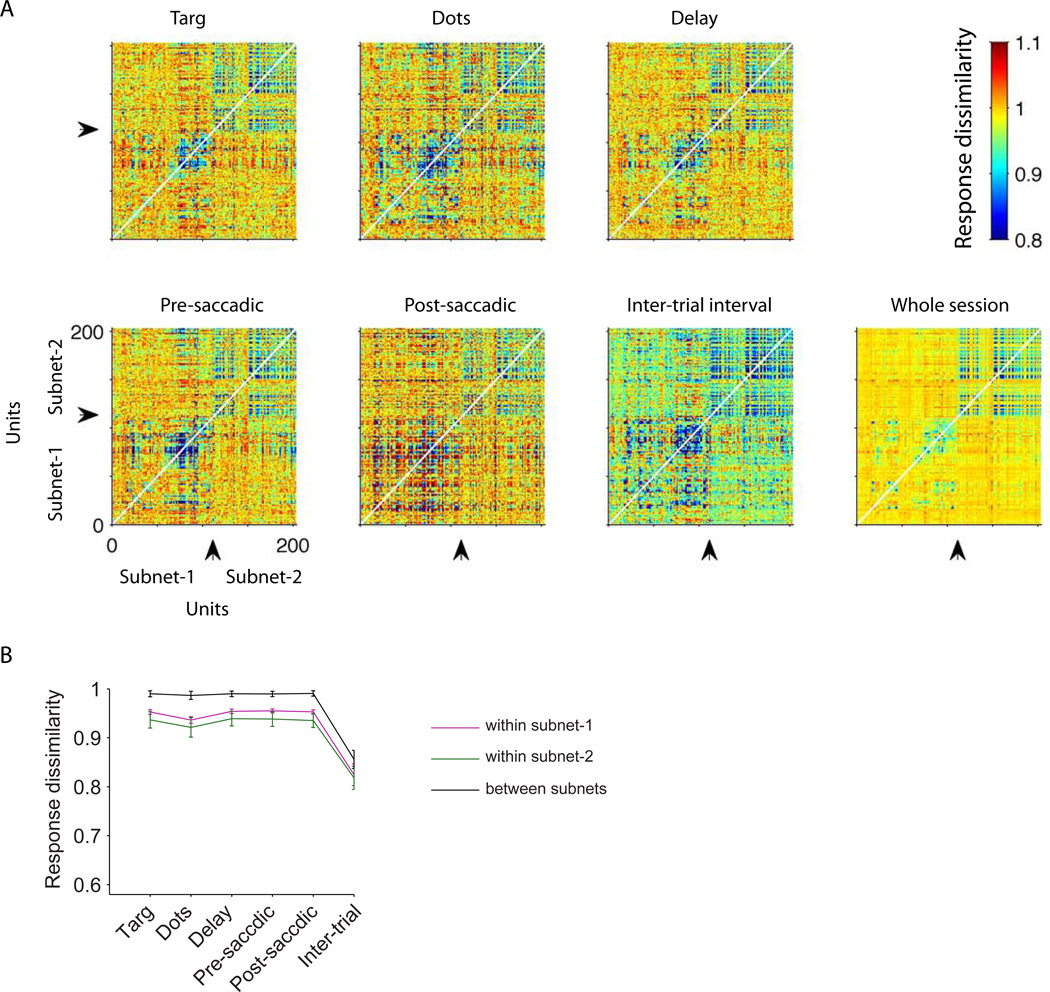
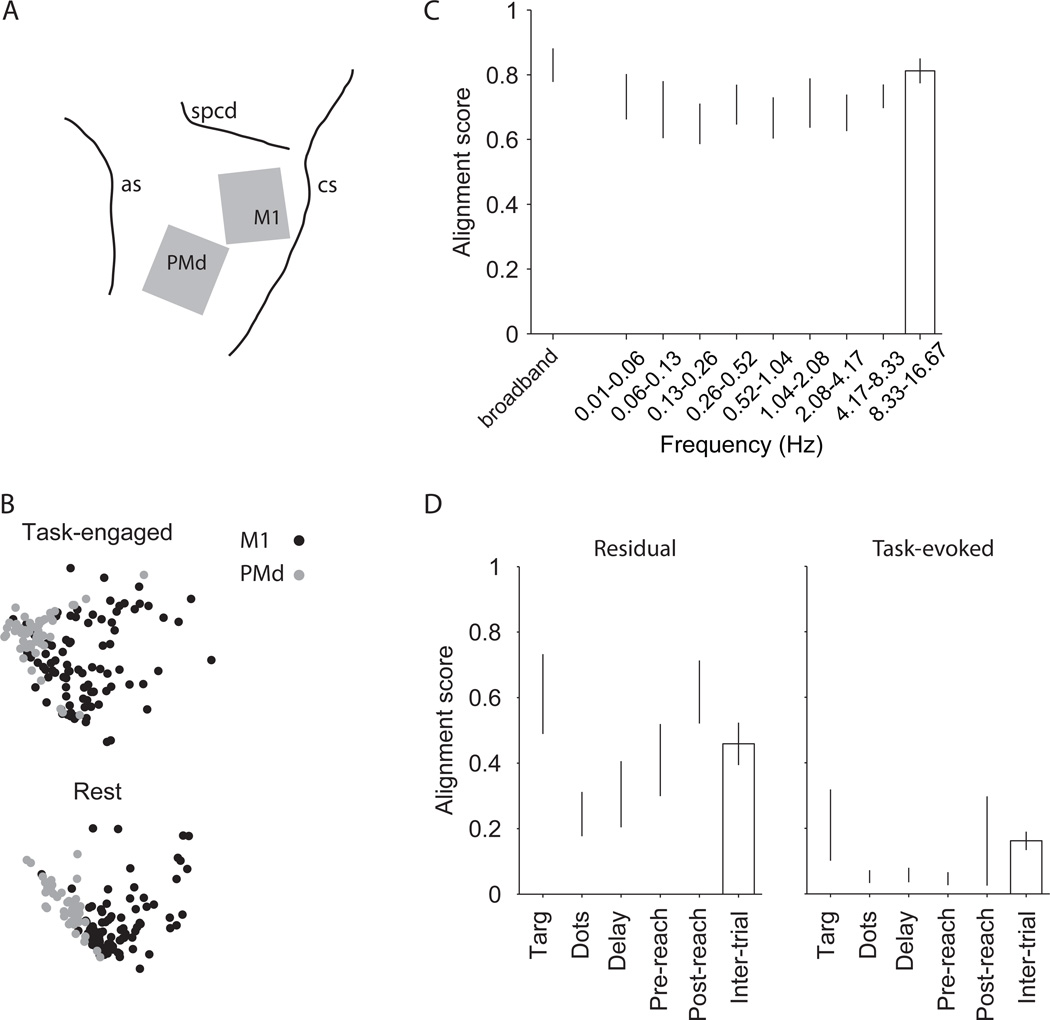
Comment in
-
Noisy neurons, neat networks.Neuron. 2015 Mar 18;85(6):1155-7. doi: 10.1016/j.neuron.2015.03.001. Neuron. 2015. PMID: 25789752
Similar articles
-
How not to study spontaneous activity.Neuroimage. 2009 May 1;45(4):1080-9. doi: 10.1016/j.neuroimage.2009.01.010. Neuroimage. 2009. PMID: 19344685
-
Mapping the mouse brain with rs-fMRI: An optimized pipeline for functional network identification.Neuroimage. 2015 Dec;123:11-21. doi: 10.1016/j.neuroimage.2015.07.090. Epub 2015 Aug 18. Neuroimage. 2015. PMID: 26296501
-
Detecting overlapped functional clusters in resting state fMRI with Connected Iterative Scan: a graph theory based clustering algorithm.J Neurosci Methods. 2011 Jul 15;199(1):108-18. doi: 10.1016/j.jneumeth.2011.05.001. Epub 2011 May 5. J Neurosci Methods. 2011. PMID: 21565220
-
Phase-synchronization-based parcellation of resting state fMRI signals reveals topographically organized clusters in early visual cortex.Neuroimage. 2018 Apr 15;170:424-433. doi: 10.1016/j.neuroimage.2017.08.063. Epub 2017 Sep 1. Neuroimage. 2018. PMID: 28867341 Review.
-
Monkey cortex through fMRI glasses.Neuron. 2014 Aug 6;83(3):533-50. doi: 10.1016/j.neuron.2014.07.015. Neuron. 2014. PMID: 25102559 Free PMC article. Review.
Cited by
-
Stabilization of a brain-computer interface via the alignment of low-dimensional spaces of neural activity.Nat Biomed Eng. 2020 Jul;4(7):672-685. doi: 10.1038/s41551-020-0542-9. Epub 2020 Apr 20. Nat Biomed Eng. 2020. PMID: 32313100 Free PMC article.
-
Nonmonotonic spatial structure of interneuronal correlations in prefrontal microcircuits.Proc Natl Acad Sci U S A. 2018 Apr 10;115(15):E3539-E3548. doi: 10.1073/pnas.1802356115. Epub 2018 Mar 27. Proc Natl Acad Sci U S A. 2018. PMID: 29588415 Free PMC article.
-
Transitions between Multiband Oscillatory Patterns Characterize Memory-Guided Perceptual Decisions in Prefrontal Circuits.J Neurosci. 2016 Jan 13;36(2):489-505. doi: 10.1523/JNEUROSCI.3678-15.2016. J Neurosci. 2016. PMID: 26758840 Free PMC article.
-
Role of Prefrontal Persistent Activity in Working Memory.Front Syst Neurosci. 2016 Jan 5;9:181. doi: 10.3389/fnsys.2015.00181. eCollection 2015. Front Syst Neurosci. 2016. PMID: 26778980 Free PMC article. Review.
-
Statistical methods for dissecting interactions between brain areas.Curr Opin Neurobiol. 2020 Dec;65:59-69. doi: 10.1016/j.conb.2020.09.009. Epub 2020 Nov 1. Curr Opin Neurobiol. 2020. PMID: 33142111 Free PMC article. Review.
References
-
- Amir Y, Harel M, Malach R. Cortical hierarchy reflected in the organization of intrinsic connections in macaque monkey visual cortex. J Comp Neurol. 1993;334:19–46. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
