

Heparanase activates the syndecan-syntenin-ALIX exosome pathway
- PMID: 25732677
- PMCID: PMC4387558
- DOI: 10.1038/cr.2015.29
Heparanase activates the syndecan-syntenin-ALIX exosome pathway
Abstract
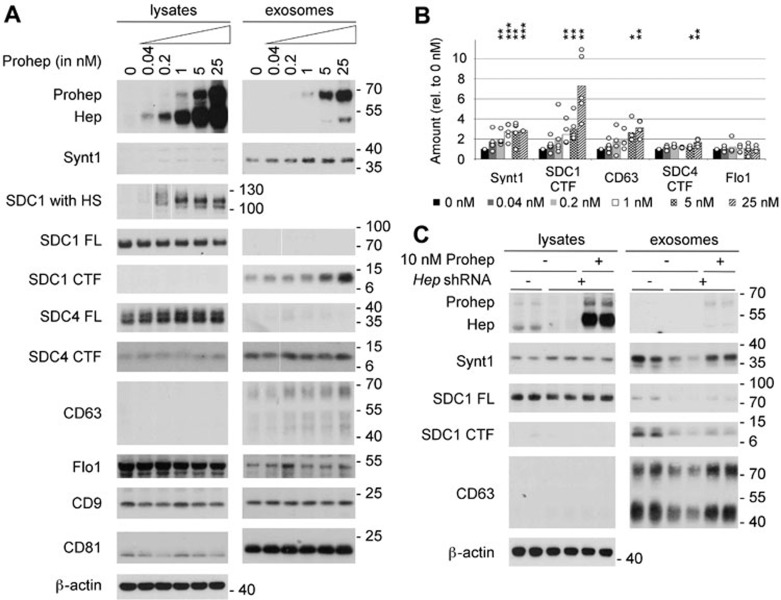
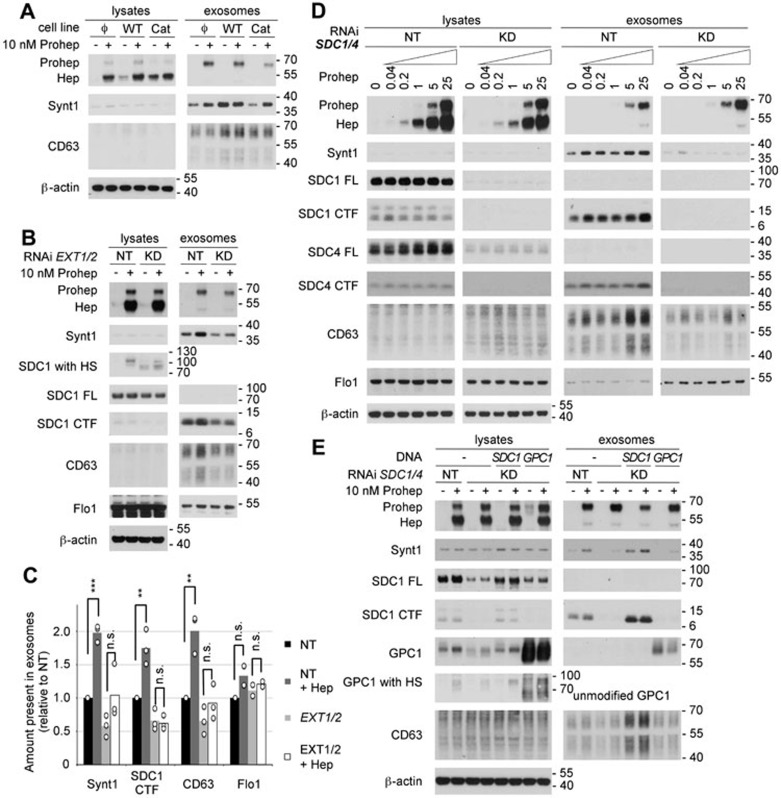
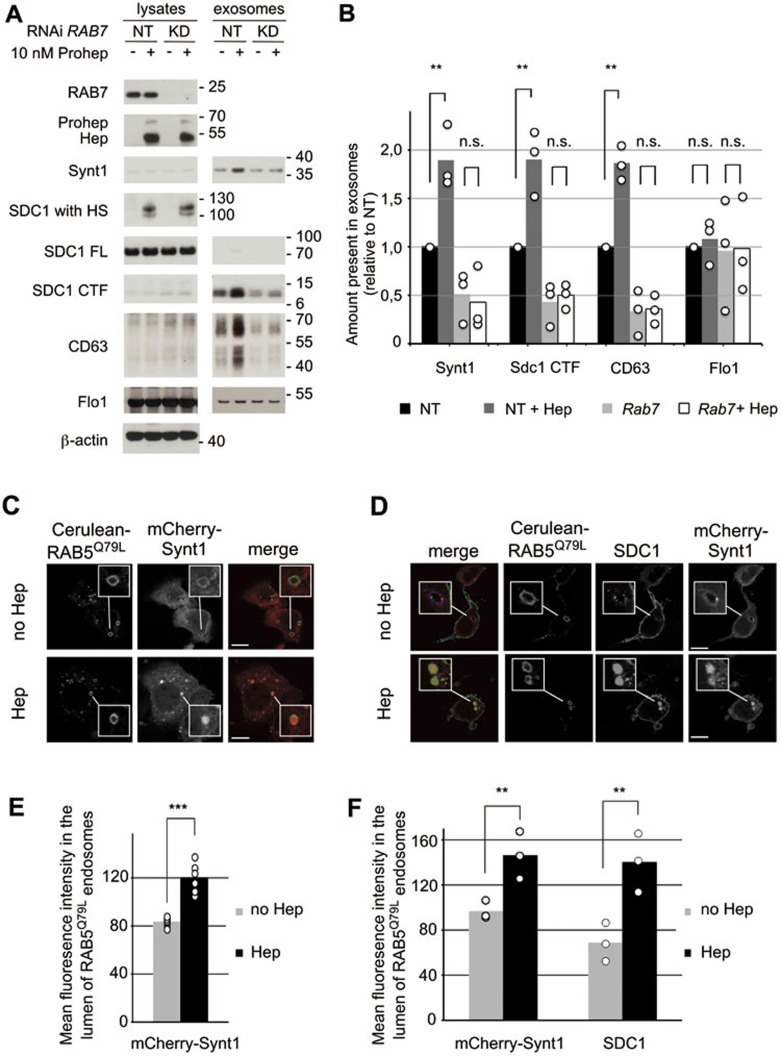
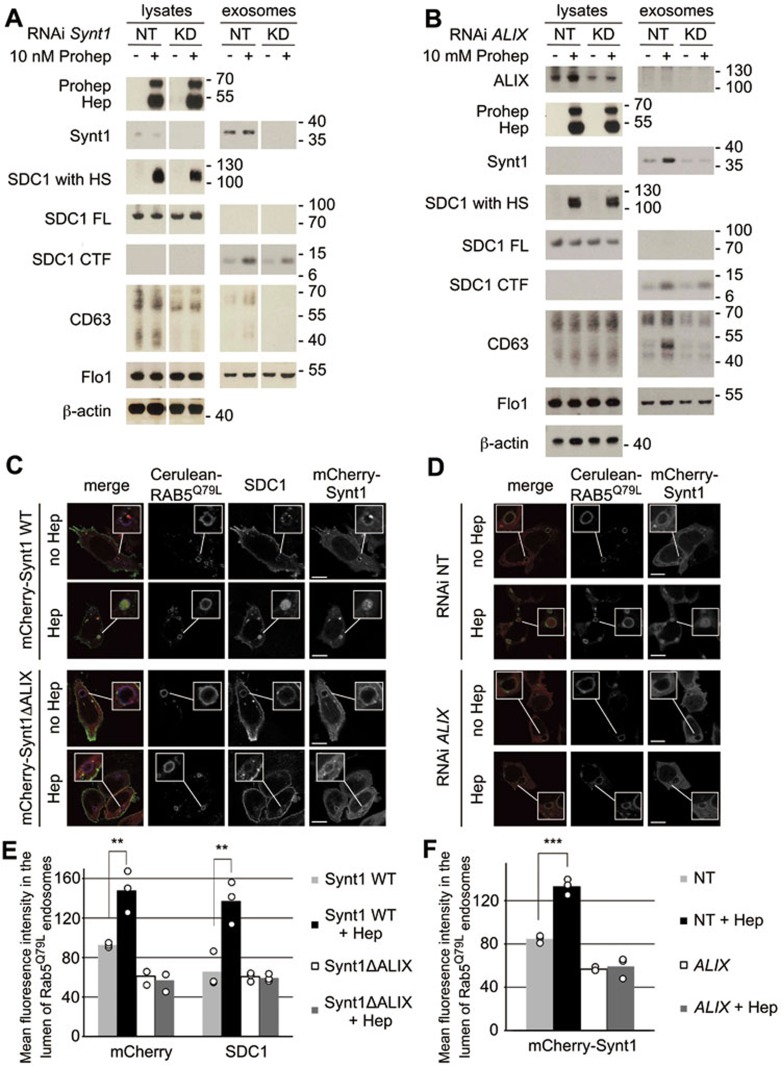
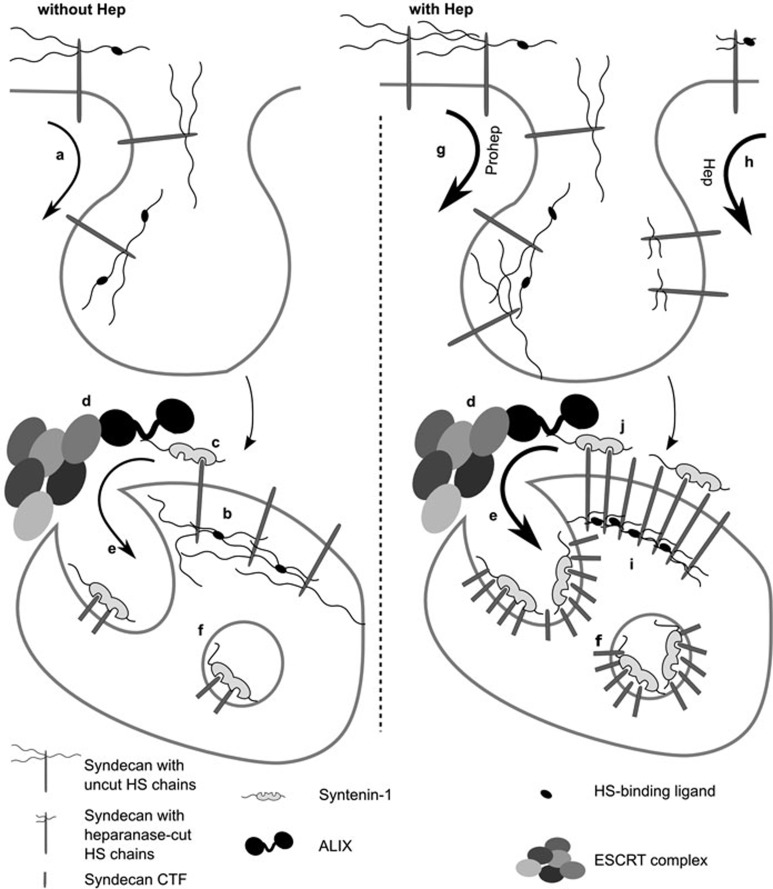
Exosomes are secreted vesicles of endosomal origin involved in signaling processes. We recently showed that the syndecan heparan sulfate proteoglycans control the biogenesis of exosomes through their interaction with syntenin-1 and the endosomal-sorting complex required for transport accessory component ALIX. Here we investigated the role of heparanase, the only mammalian enzyme able to cleave heparan sulfate internally, in the syndecan-syntenin-ALIX exosome biogenesis pathway. We show that heparanase stimulates the exosomal secretion of syntenin-1, syndecan and certain other exosomal cargo, such as CD63, in a concentration-dependent manner. In contrast, exosomal CD9, CD81 and flotillin-1 are not affected. Conversely, reduction of endogenous heparanase reduces the secretion of syntenin-1-containing exosomes. The ability of heparanase to stimulate exosome production depends on syntenin-1 and ALIX. Syndecans, but not glypicans, support exosome biogenesis in heparanase-exposed cells. Finally, heparanase stimulates intraluminal budding of syndecan and syntenin-1 in endosomes, depending on the syntenin-ALIX interaction. Taken together, our findings identify heparanase as a modulator of the syndecan-syntenin-ALIX pathway, fostering endosomal membrane budding and the biogenesis of exosomes by trimming the heparan sulfate chains on syndecans. In addition, our data suggest that this mechanism controls the selection of specific cargo to exosomes.
Figures
Comment in
-
Resolving sorting mechanisms into exosomes.Cell Res. 2015 May;25(5):531-2. doi: 10.1038/cr.2015.39. Epub 2015 Mar 31. Cell Res. 2015. PMID: 25828531 Free PMC article.
References
-
- Valadi H, Ekstrom K, Bossios A, Sjostrand M, Lee JJ, Lotvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol. 2007;9:654–659. - PubMed
-
- Thery C, Duban L, Segura E, Veron P, Lantz O, Amigorena S. Indirect activation of naive CD4+ T cells by dendritic cell-derived exosomes. Nat Immunol. 2002;3:1156–1162. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
