

Sociality in Escherichia coli: Enterochelin Is a Private Good at Low Cell Density and Can Be Shared at High Cell Density
- PMID: 25733620
- PMCID: PMC4455259
- DOI: 10.1128/JB.02596-14
Sociality in Escherichia coli: Enterochelin Is a Private Good at Low Cell Density and Can Be Shared at High Cell Density
Abstract
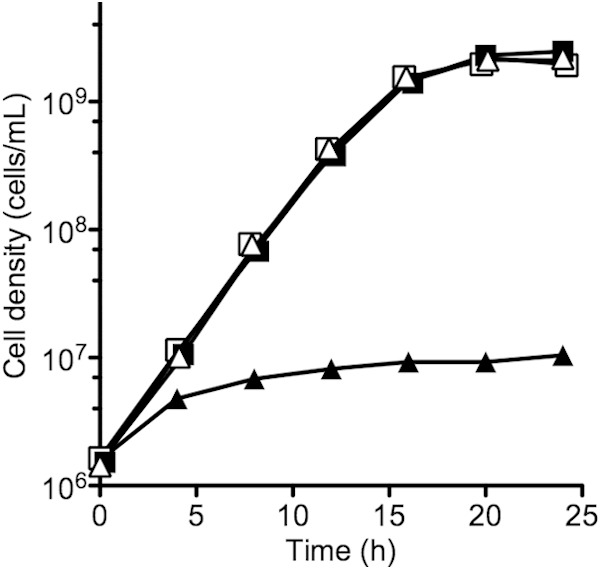
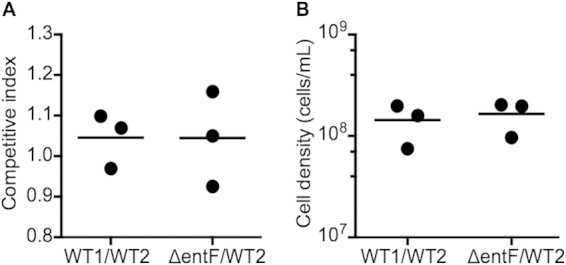
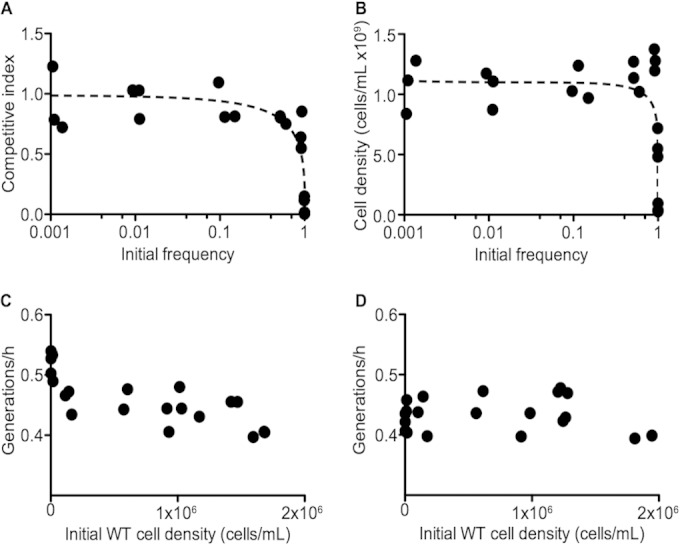
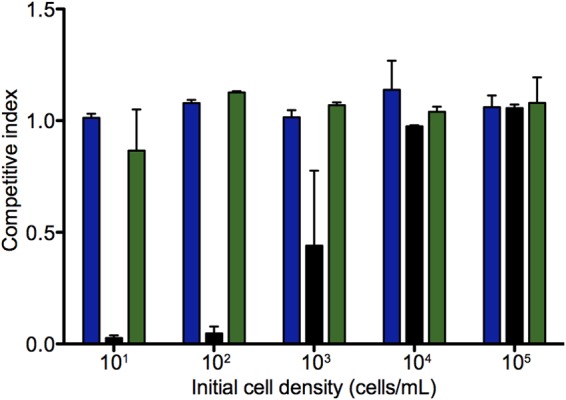
Many bacteria produce secreted iron chelators called siderophores, which can be shared among cells with specific siderophore uptake systems regardless of whether the cell produces siderophores. Sharing secreted products allows freeloading, where individuals use resources without bearing the cost of production. Here we show that the Escherichia coli siderophore enterochelin is not evenly shared between producers and nonproducers. Wild-type Escherichia coli grows well in low-iron minimal medium, and an isogenic enterochelin synthesis mutant (ΔentF) grows very poorly. The enterochelin mutant grows well in low-iron medium supplemented with enterochelin. At high cell densities the ΔentF mutant can compete equally with the wild type in low-iron medium. At low cell densities the ΔentF mutant cannot compete. Furthermore, the growth rate of the wild type is unaffected by cell density. The wild type grows well in low-iron medium even at very low starting densities. Our experiments support a model where at least some enterochelin remains associated with the cells that produce it, and the cell-associated enterochelin enables iron acquisition even at very low cell density. Enterochelin that is not retained by producing cells at low density is lost to dilution. At high cell densities, cell-free enterochelin can accumulate and be shared by all cells in the group. Partial privatization is a solution to the problem of iron acquisition in low-iron, low-cell-density habitats. Cell-free enterochelin allows for iron scavenging at a distance at higher population densities. Our findings shed light on the conditions under which freeloaders might benefit from enterochelin uptake systems.
Importance: Sociality in microbes has become a topic of great interest. One facet of sociality is the sharing of secreted products, such as the iron-scavenging siderophores. We present evidence that the Escherichia coli siderophore enterochelin is relatively inexpensive to produce and is partially privatized such that it can be efficiently shared only at high producer cell densities. At low cell densities, cell-free enterochelin is scarce and only enterochelin producers are able to grow in low-iron medium. Because freely shared products can be exploited by freeloaders, this partial privatization may help explain how enterochelin production is stabilized in E. coli and may provide insight into when enterochelin is available for freeloaders.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Neilands JB, Konopka K, Schwyn B, Coy M, Francis RT, Paw BH, Bagg A. 1987. Comparative biochemistry of microbial iron assimilation, p 3–33. In Winkelmann G, Van der Helm D, Neilands JB (ed), Iron transport in microbes, plants, and animals. VCH, Weinheim, Federal Republic of Germany.
-
- Lankford CE, Byers BR. 1973. Bacterial assimilation of iron. Crit Rev Microbiol 2:273–331. doi:10.3109/10408417309108388. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
