

MinD-like ATPase FlhG effects location and number of bacterial flagella during C-ring assembly
- PMID: 25733861
- PMCID: PMC4364217
- DOI: 10.1073/pnas.1419388112
MinD-like ATPase FlhG effects location and number of bacterial flagella during C-ring assembly
Abstract
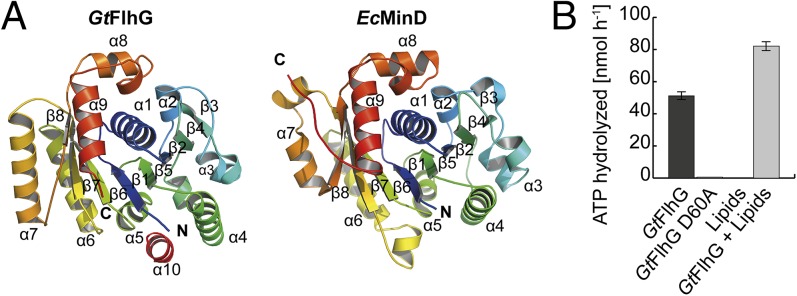
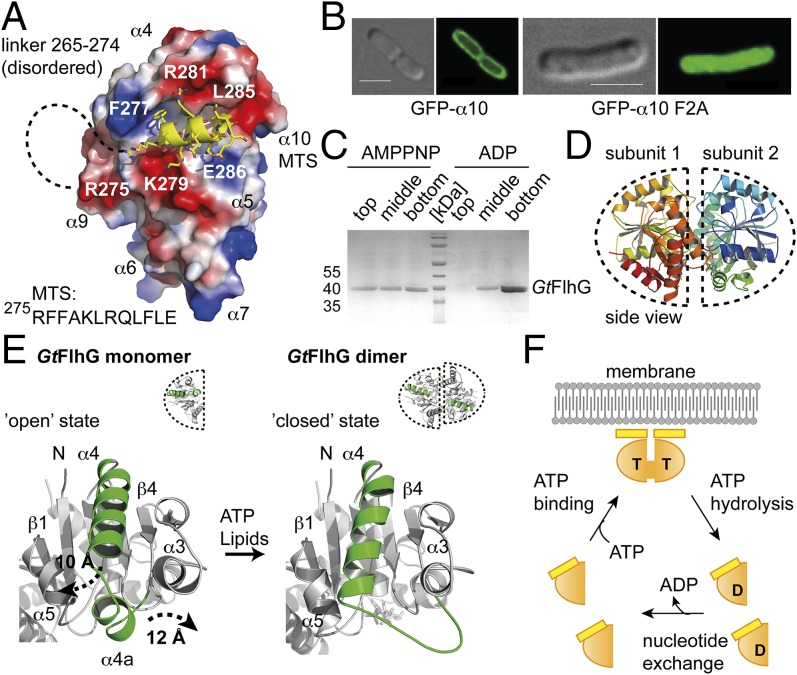
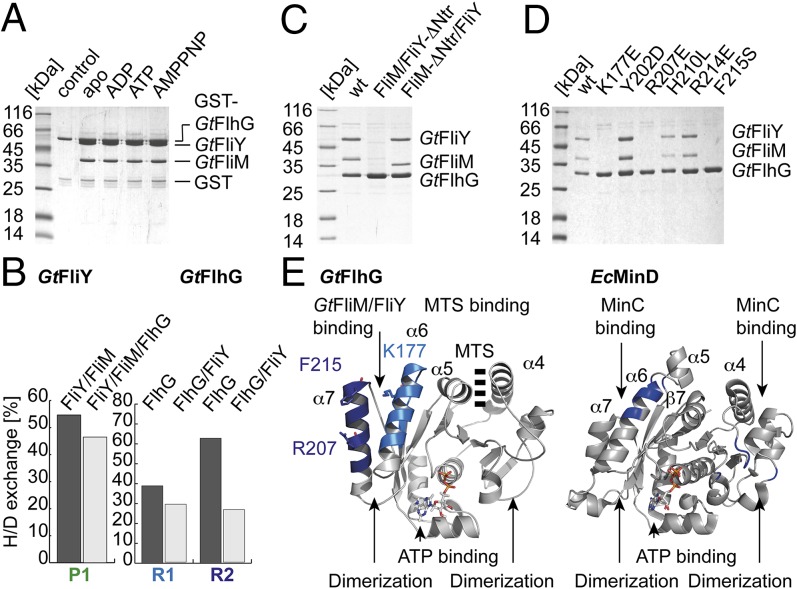
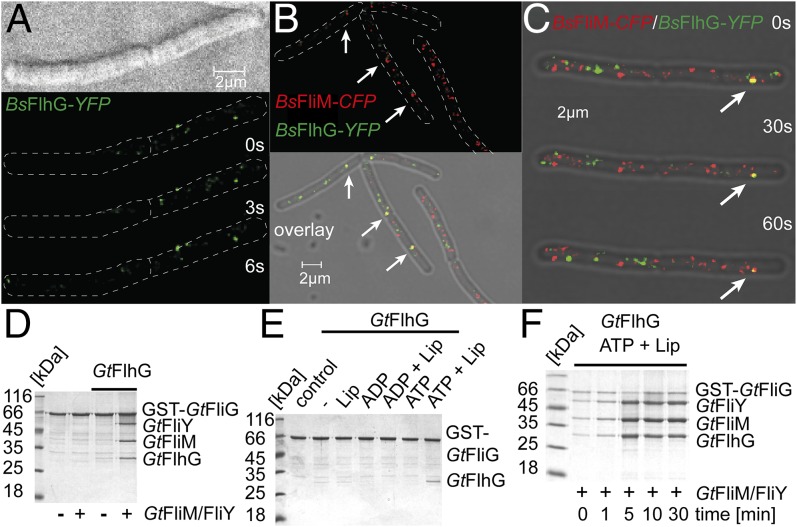
The number and location of flagella, bacterial organelles of locomotion, are species specific and appear in regular patterns that represent one of the earliest taxonomic criteria in microbiology. However, the mechanisms that reproducibly establish these patterns during each round of cell division are poorly understood. FlhG (previously YlxH) is a major determinant for a variety of flagellation patterns. Here, we show that FlhG is a structural homolog of the ATPase MinD, which serves in cell-division site determination. Like MinD, FlhG forms homodimers that are dependent on ATP and lipids. It interacts with a complex of the flagellar C-ring proteins FliM and FliY (also FliN) in the Gram-positive, peritrichous-flagellated Bacillus subtilis and the Gram-negative, polar-flagellated Shewanella putrefaciens. FlhG interacts with FliM/FliY in a nucleotide-independent manner and activates FliM/FliY to assemble with the C-ring protein FliG in vitro. FlhG-driven assembly of the FliM/FliY/FliG complex is strongly enhanced by ATP and lipids. The protein shows a highly dynamic subcellular distribution between cytoplasm and flagellar basal bodies, suggesting that FlhG effects flagellar location and number during assembly of the C-ring. We describe the molecular evolution of a MinD-like ATPase into a flagellation pattern effector and suggest that the underappreciated structural diversity of the C-ring proteins might contribute to the formation of different flagellation patterns.
Keywords: Bacillus; C-ring; FlhG; Shewanella; flagellum.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Altegoer F, Schuhmacher J, Pausch P, Bange G. From molecular evolution to biobricks and synthetic modules: A lesson by the bacterial flagellum. Biotechnol Genet Eng Rev. 2014;30:49–64. - PubMed
-
- Kusumoto A, et al. Regulation of polar flagellar number by the flhF and flhG genes in Vibrio alginolyticus. J Biochem. 2006;139(1):113–121. - PubMed
-
- Campos-García J, Nájera R, Camarena L, Soberón-Chávez G. The pseudomonas aeruginosa motR gene involved in regulation of bacterial motility. FEMS Microbiol Lett. 2000;184(1):57–62. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
