

Single-cell analysis reveals transcriptional heterogeneity of neural progenitors in human cortex
- PMID: 25734491
- PMCID: PMC5568903
- DOI: 10.1038/nn.3980
Single-cell analysis reveals transcriptional heterogeneity of neural progenitors in human cortex
Abstract
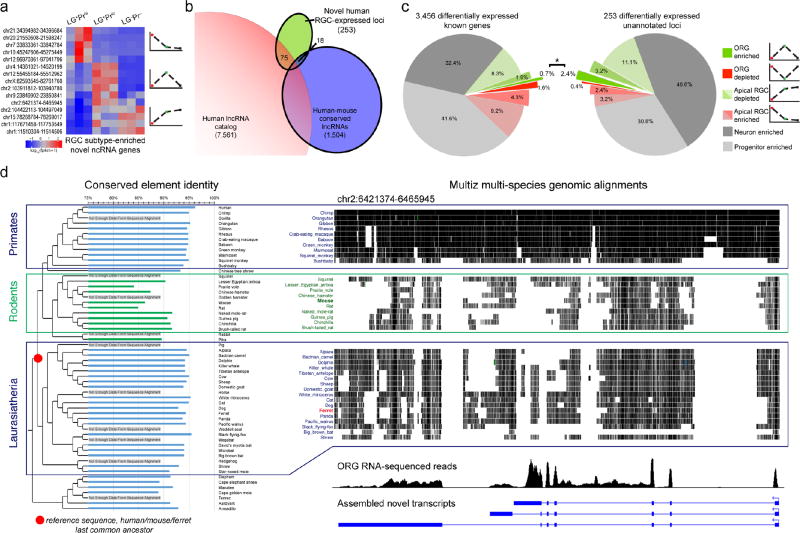
The human cerebral cortex depends for its normal development and size on a precisely controlled balance between self-renewal and differentiation of diverse neural progenitor cells. Specialized progenitors that are common in humans but virtually absent in rodents, called outer radial glia (ORG), have been suggested to be crucial to the evolutionary expansion of the human cortex. We combined progenitor subtype-specific sorting with transcriptome-wide RNA sequencing to identify genes enriched in human ORG, which included targets of the transcription factor neurogenin and previously uncharacterized, evolutionarily dynamic long noncoding RNAs. Activating the neurogenin pathway in ferret progenitors promoted delamination and outward migration. Finally, single-cell transcriptional profiling in human, ferret and mouse revealed more cells coexpressing proneural neurogenin targets in human than in other species, suggesting greater neuronal lineage commitment and differentiation of self-renewing progenitors. Thus, we find that the abundance of human ORG is paralleled by increased transcriptional heterogeneity of cortical progenitors.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
