

PRMT9 is a type II methyltransferase that methylates the splicing factor SAP145
- PMID: 25737013
- PMCID: PMC4351962
- DOI: 10.1038/ncomms7428
PRMT9 is a type II methyltransferase that methylates the splicing factor SAP145
Abstract
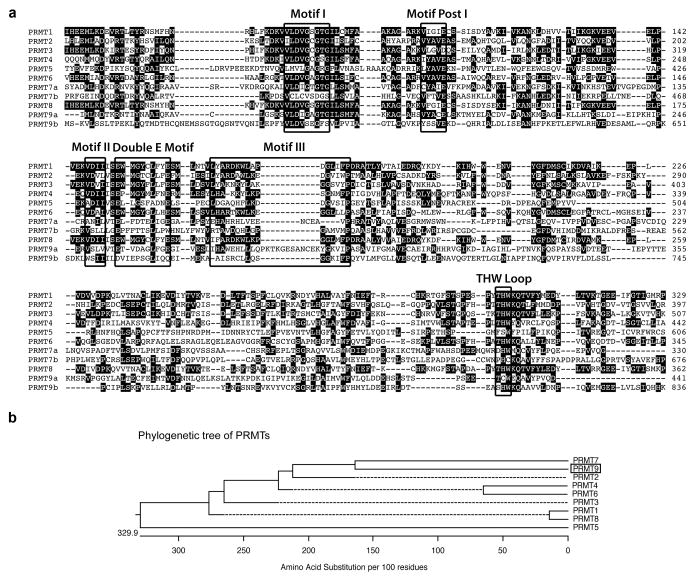
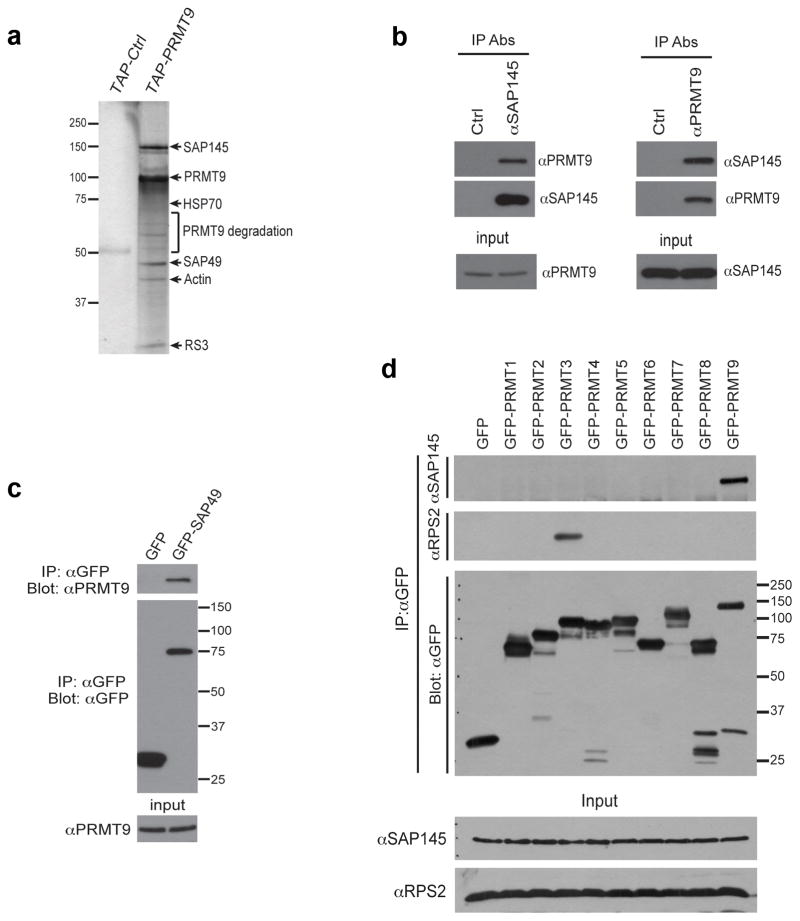
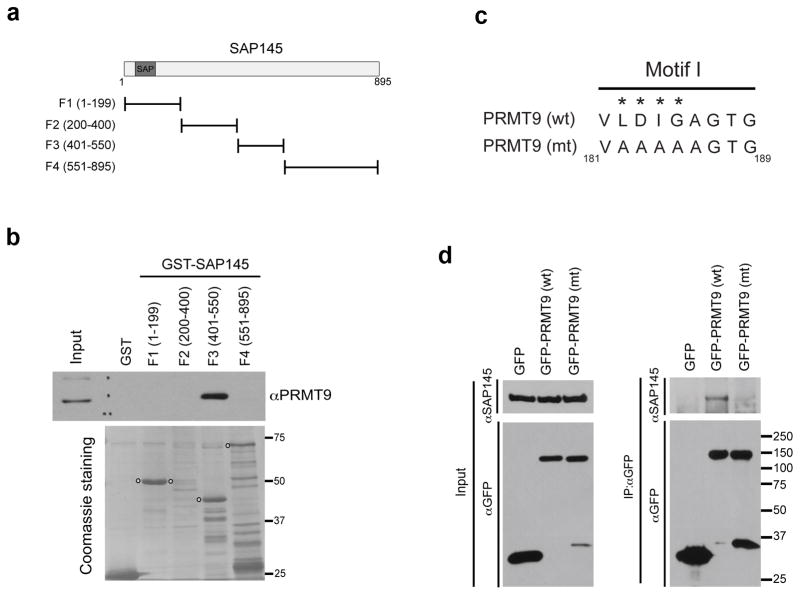
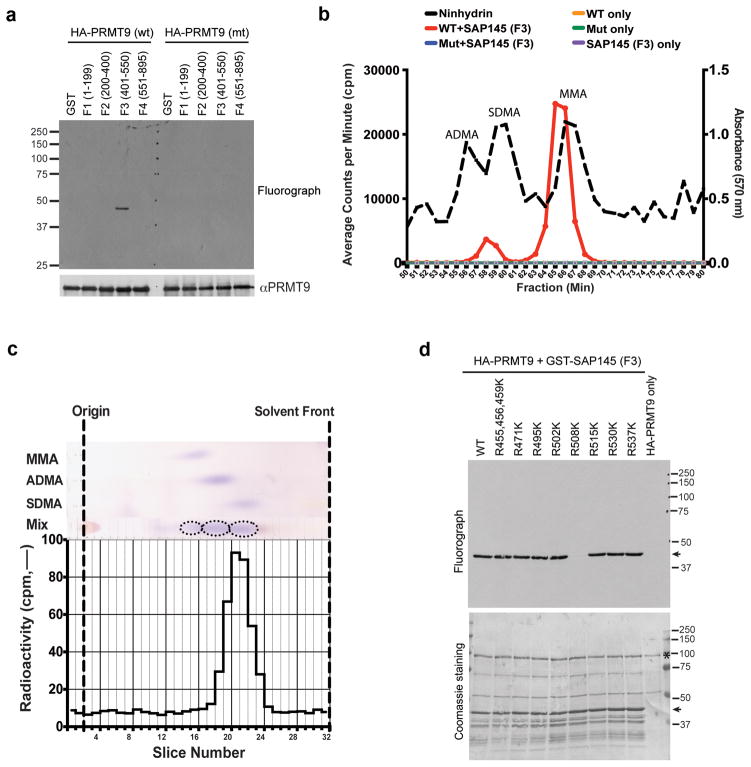
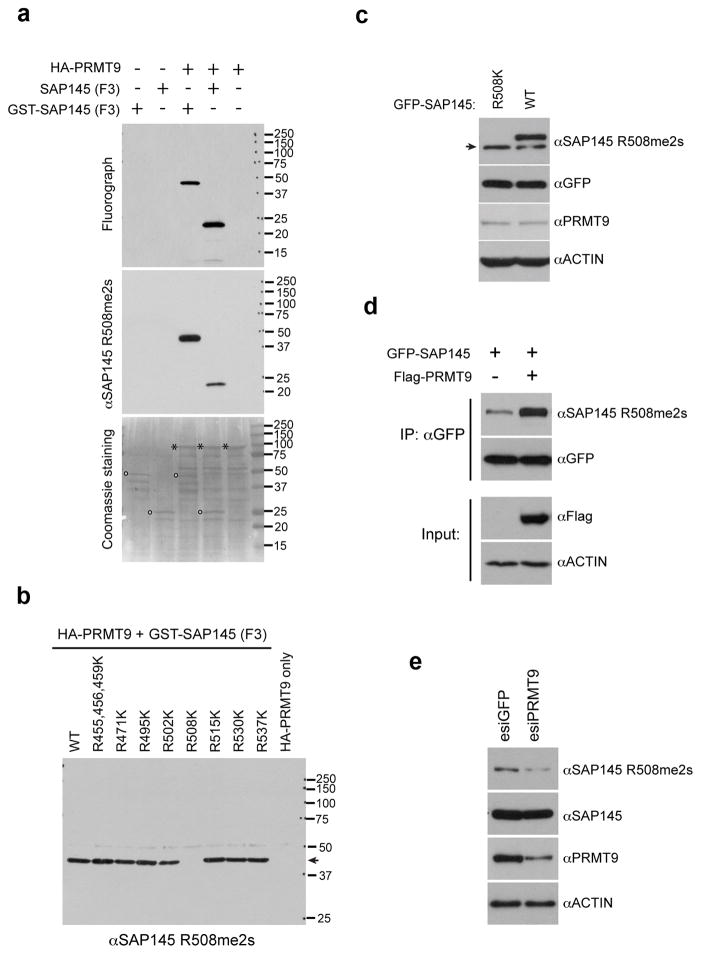
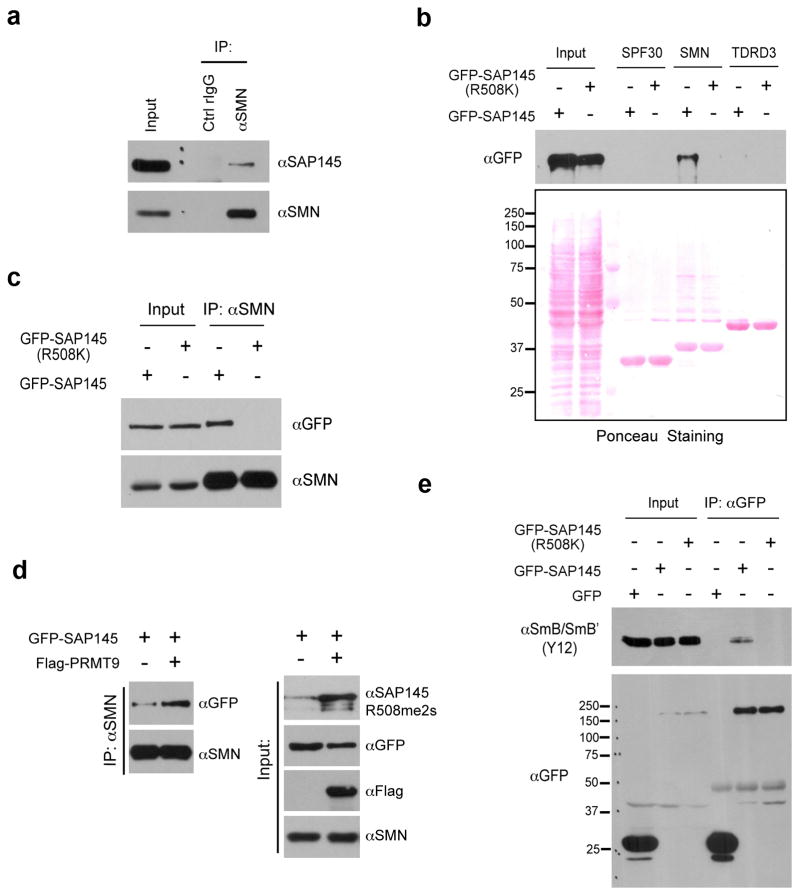
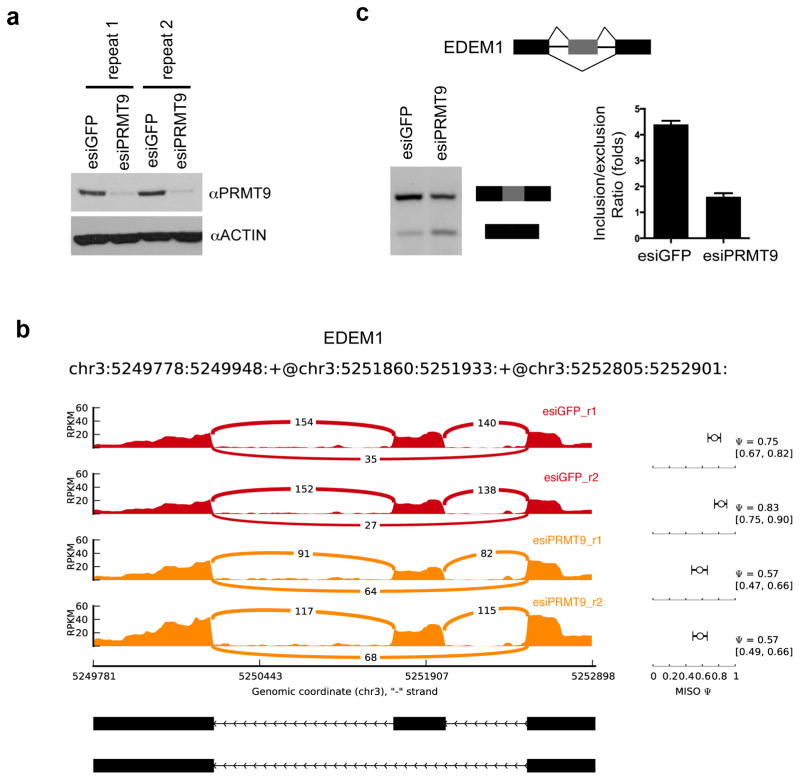
The human genome encodes a family of nine protein arginine methyltransferases (PRMT1-9), whose members can catalyse three distinct types of methylation on arginine residues. Here we identify two spliceosome-associated proteins-SAP145 and SAP49-as PRMT9-binding partners, linking PRMT9 to U2 snRNP maturation. We show that SAP145 is methylated by PRMT9 at arginine 508, which takes the form of monomethylated arginine (MMA) and symmetrically dimethylated arginine (SDMA). PRMT9 thus joins PRMT5 as the only mammalian enzymes capable of depositing the SDMA mark. Methylation of SAP145 on Arg 508 generates a binding site for the Tudor domain of the Survival of Motor Neuron (SMN) protein, and RNA-seq analysis reveals gross splicing changes when PRMT9 levels are attenuated. These results identify PRMT9 as a nonhistone methyltransferase that primes the U2 snRNP for interaction with SMN.
Conflict of interest statement
Figures
Similar articles
-
Unique Features of Human Protein Arginine Methyltransferase 9 (PRMT9) and Its Substrate RNA Splicing Factor SF3B2.J Biol Chem. 2015 Jul 3;290(27):16723-43. doi: 10.1074/jbc.M115.659433. Epub 2015 May 15. J Biol Chem. 2015. PMID: 25979344 Free PMC article.
-
FBXO11/PRMT9, a new protein arginine methyltransferase, symmetrically dimethylates arginine residues.Biochem Biophys Res Commun. 2006 Apr 7;342(2):472-81. doi: 10.1016/j.bbrc.2006.01.167. Epub 2006 Feb 8. Biochem Biophys Res Commun. 2006. PMID: 16487488
-
Loss-of-function mutation in PRMT9 causes abnormal synapse development by dysregulation of RNA alternative splicing.Nat Commun. 2024 Apr 1;15(1):2809. doi: 10.1038/s41467-024-47107-9. Nat Commun. 2024. PMID: 38561334 Free PMC article.
-
Chemical probes for protein arginine methyltransferases.Methods. 2020 Mar 15;175:30-43. doi: 10.1016/j.ymeth.2019.11.017. Epub 2019 Dec 3. Methods. 2020. PMID: 31809836 Review.
-
Protein arginine methylation: Cellular functions and methods of analysis.Biochim Biophys Acta. 2006 Dec;1764(12):1890-903. doi: 10.1016/j.bbapap.2006.08.008. Epub 2006 Aug 25. Biochim Biophys Acta. 2006. PMID: 17010682 Review.
Cited by
-
Uncovering epigenetic landscape: a new path for biomarkers identification and drug development.Mol Biol Rep. 2020 Nov;47(11):9097-9122. doi: 10.1007/s11033-020-05916-3. Epub 2020 Oct 21. Mol Biol Rep. 2020. PMID: 33089404 Review.
-
Closing in on human methylation-the versatile family of seven-β-strand (METTL) methyltransferases.Nucleic Acids Res. 2024 Oct 28;52(19):11423-11441. doi: 10.1093/nar/gkae816. Nucleic Acids Res. 2024. PMID: 39351878 Free PMC article. Review.
-
Mouse Models of Overexpression Reveal Distinct Oncogenic Roles for Different Type I Protein Arginine Methyltransferases.Cancer Res. 2019 Jan 1;79(1):21-32. doi: 10.1158/0008-5472.CAN-18-1995. Epub 2018 Oct 23. Cancer Res. 2019. PMID: 30352814 Free PMC article.
-
Developmental expression of three prmt genes in Xenopus.Zool Res. 2019 Mar 18;40(2):102-107. doi: 10.24272/j.issn.2095-8137.2018.064. Epub 2018 Aug 20. Zool Res. 2019. PMID: 30127333 Free PMC article.
-
Caenorhabditis elegans PRMT-7 and PRMT-9 Are Evolutionarily Conserved Protein Arginine Methyltransferases with Distinct Substrate Specificities.Biochemistry. 2017 May 23;56(20):2612-2626. doi: 10.1021/acs.biochem.7b00283. Epub 2017 May 9. Biochemistry. 2017. PMID: 28441492 Free PMC article.
References
-
- Gary JD, Clarke S. RNA and protein interactions modulated by protein arginine methylation. Prog Nucleic Acid Res Mol Biol. 1998;61:65–131. - PubMed
-
- Boffa LC, Karn J, Vidali G, Allfrey VG. Distribution of NG, NG,-dimethylarginine in nuclear protein fractions. Biochem Biophys Res Commun. 1977;74:969–976. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
