

Cutaneous Na+ storage strengthens the antimicrobial barrier function of the skin and boosts macrophage-driven host defense
- PMID: 25738463
- PMCID: PMC4350016
- DOI: 10.1016/j.cmet.2015.02.003
Cutaneous Na+ storage strengthens the antimicrobial barrier function of the skin and boosts macrophage-driven host defense
Abstract
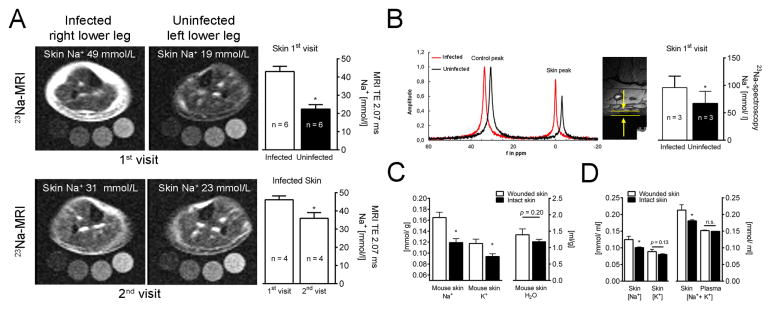
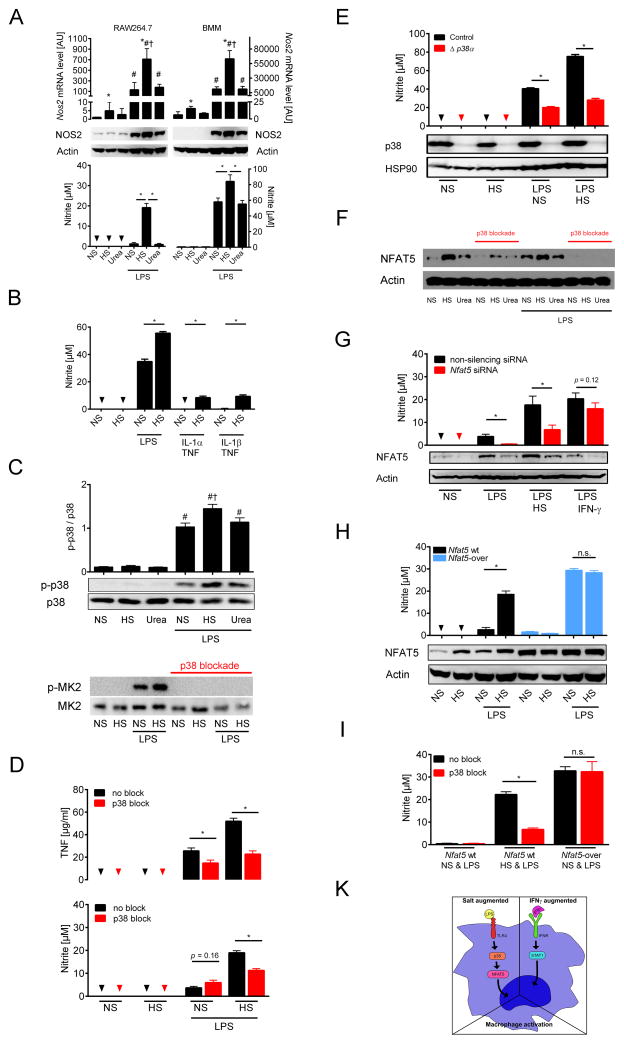
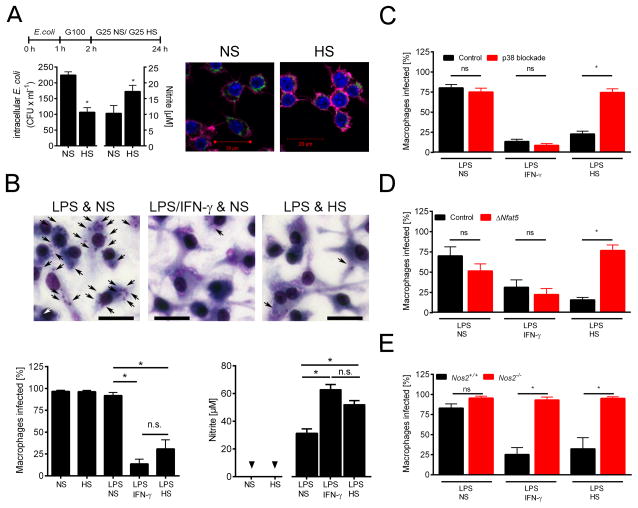
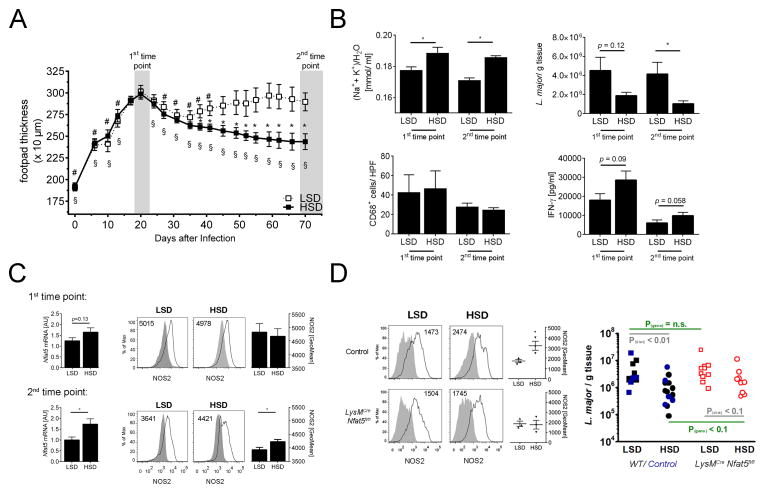
Immune cells regulate a hypertonic microenvironment in the skin; however, the biological advantage of increased skin Na(+) concentrations is unknown. We found that Na(+) accumulated at the site of bacterial skin infections in humans and in mice. We used the protozoan parasite Leishmania major as a model of skin-prone macrophage infection to test the hypothesis that skin-Na(+) storage facilitates antimicrobial host defense. Activation of macrophages in the presence of high NaCl concentrations modified epigenetic markers and enhanced p38 mitogen-activated protein kinase (p38/MAPK)-dependent nuclear factor of activated T cells 5 (NFAT5) activation. This high-salt response resulted in elevated type-2 nitric oxide synthase (Nos2)-dependent NO production and improved Leishmania major control. Finally, we found that increasing Na(+) content in the skin by a high-salt diet boosted activation of macrophages in a Nfat5-dependent manner and promoted cutaneous antimicrobial defense. We suggest that the hypertonic microenvironment could serve as a barrier to infection.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Antibacterial immunity: a pinch of salt.Nat Rev Immunol. 2015 Apr;15(4):202. doi: 10.1038/nri3835. Epub 2015 Mar 13. Nat Rev Immunol. 2015. PMID: 25765202 No abstract available.
References
-
- Angrisano T, Lembo F, Peluso S, Keller S, Chiariotti L, Pero R. Helicobacter pylori regulates iNOS promoter by histone modifications in human gastric epithelial cells. Med Microbiol Immunol. 2012;201:249–257. - PubMed
-
- Belkaid Y, Segre JA. Dialogue between skin microbiota and immunity. Science. 2014;346:954–959. - PubMed
-
- Braff MH, Gallo RL. Antimicrobial peptides: an essential component of the skin defensive barrier. Current topics in microbiology and immunology. 2006;306:91–110. - PubMed
-
- Denkert C, Warskulat U, Hensel F, Haussinger D. Osmolyte strategy in human monocytes and macrophages: involvement of p38MAPK in hyperosmotic induction of betaine and myoinositol transporters. Arch Biochem Biophys. 1998;354:172–180. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
