

Dopamine D1 and D5 receptors differentially regulate oxidative stress through paraoxonase 2 in kidney cells
- PMID: 25740199
- PMCID: PMC5261865
- DOI: 10.3109/10715762.2015.1006215
Dopamine D1 and D5 receptors differentially regulate oxidative stress through paraoxonase 2 in kidney cells
Abstract
Background: The renal dopaminergic system plays an important role in the pathogenesis of hypertension. Dopamine D1-like receptors (D1R and D5R) decrease reactive oxygen species (ROS) production via inhibition of pro-oxidant enzymes such as NADPH oxidase. Paraoxonase 2 (PON2) is also involved in the inhibition of NADPH oxidase activity. Therefore, we tested the hypothesis that D1R and D5R inhibit ROS production by increasing the expression of PON2, including those in membrane microdomains.
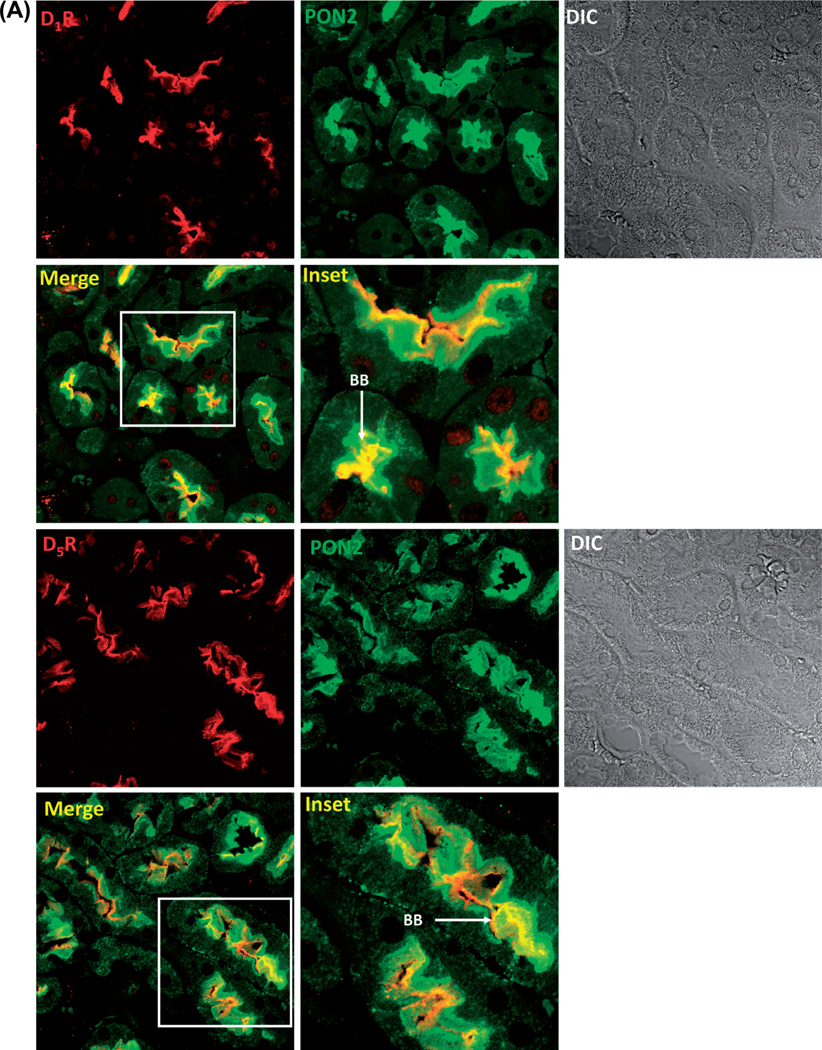
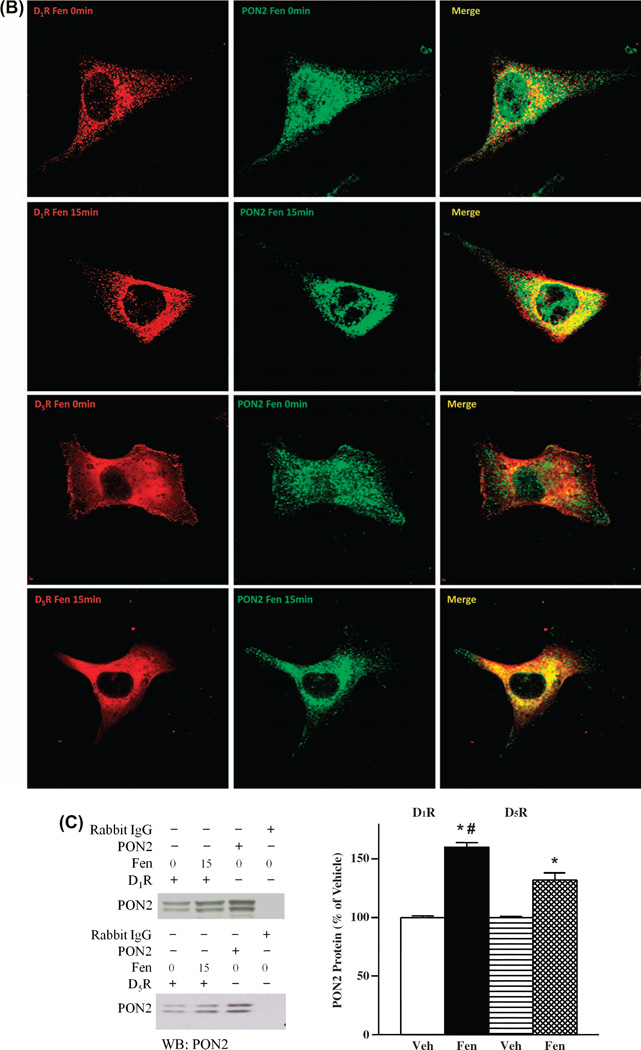
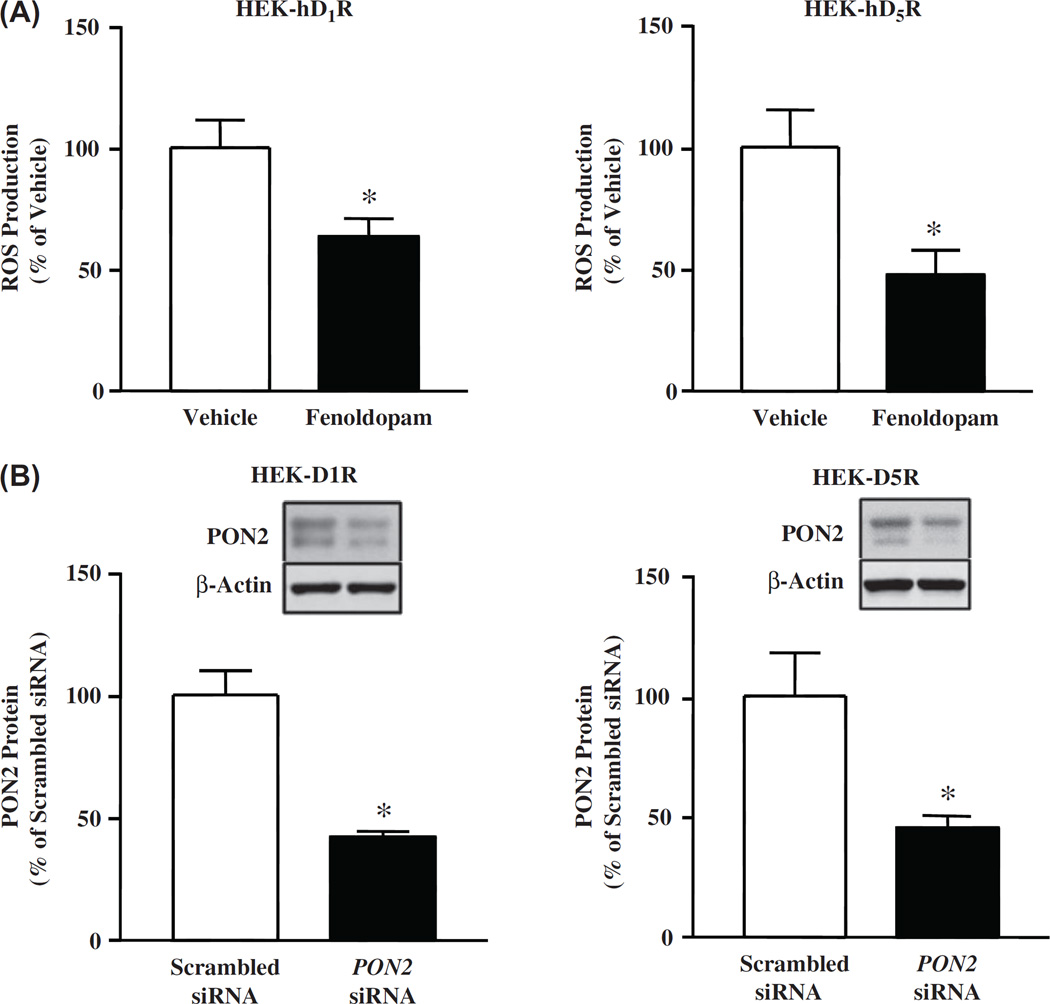
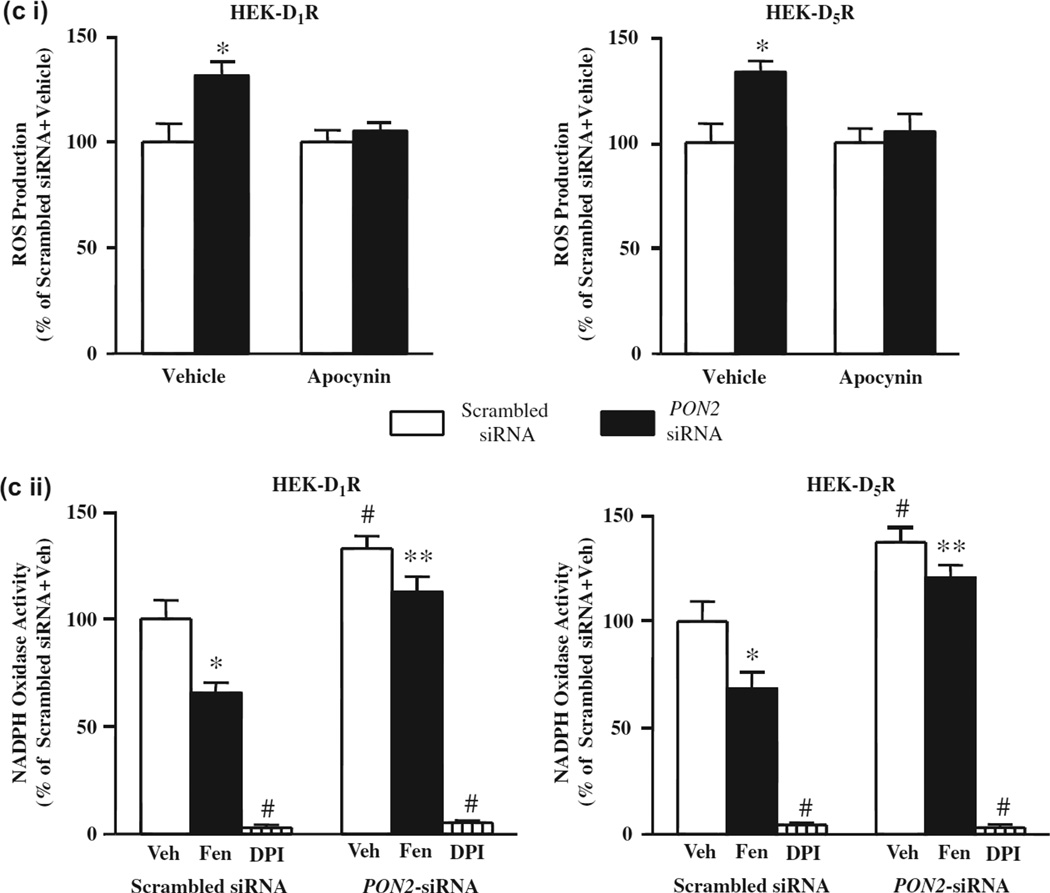
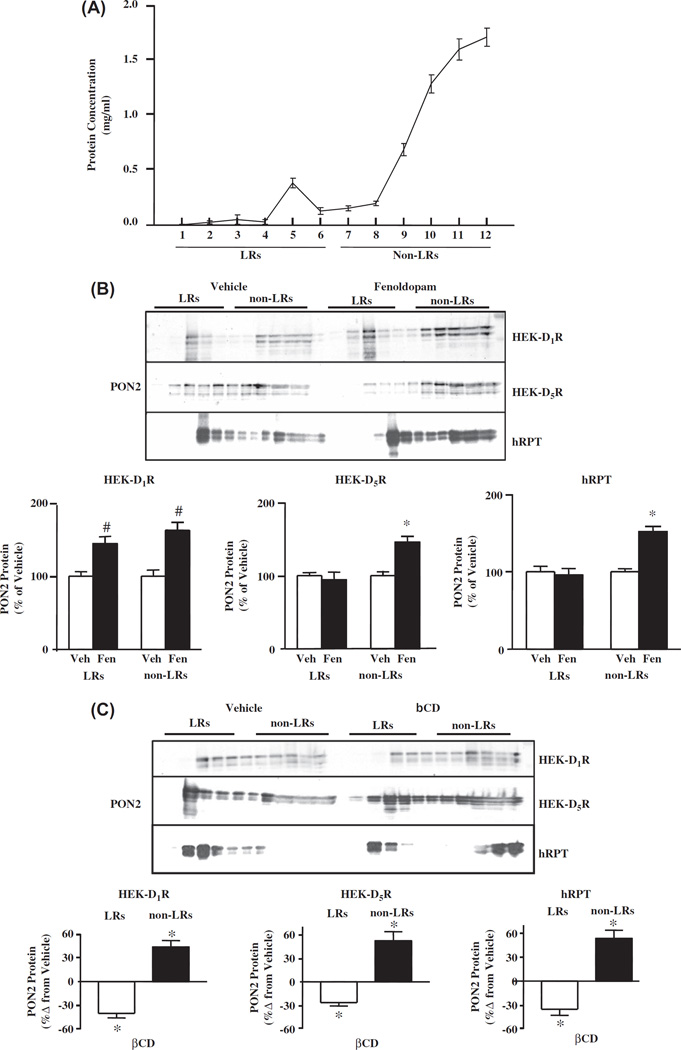
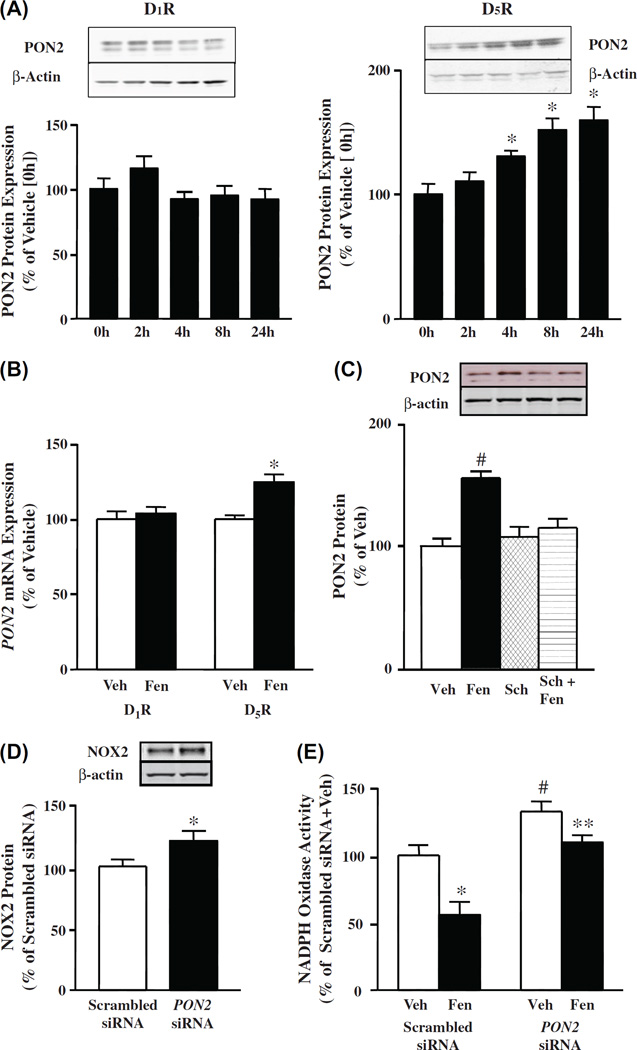
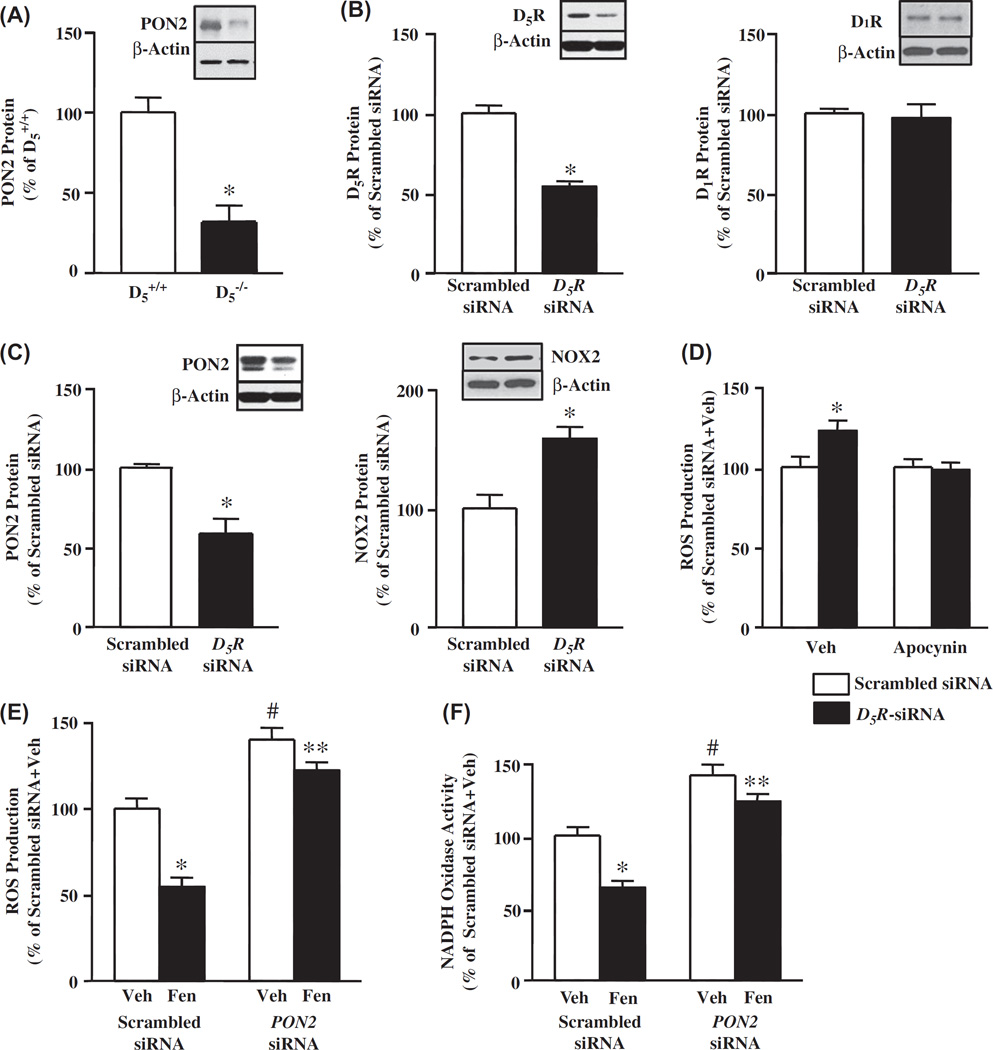
Methods and results: PON2 colocalized with D1R and D5R in mouse renal proximal tubules (RPTs), human RPT (hRPT) cells, and HEK293 cells heterologously expressing human D1R (HEK-hD1R) or D5R (HEK-hD5R). Fenoldopam, an agonist for both D1R and D5R, increased PON2 co-immunoprecipitation with D1R and D5R in HEK-hD1R and HEK-hD5R cells, respectively. Silencing PON2 increased ROS production and NADPH oxidase activity, and impaired the inhibitory effect of fenoldopam. Fenoldopam increased PON2 protein in both lipid rafts (LRs) and non-LRs in HEK-hD1R cells, but only in non-LRs in HEK-hD5R and hRPT cells. Long-term (hrs) fenoldopam stimulation increased PON2 protein in a time-dependent manner in HEK-hD5R, but not in HEK-hD1R cells. Because the effects of fenoldopam on non-LR and total PON2 expressions were similar in HEK-hD5R and hRPT cells, additional studies were performed to determine the relationship between D5R and PON2. Renal PON2 protein was decreased in D5(-/-) mice. In hRPT cells, silencing D5R decreased PON2 expression and increased ROS production.
Conclusions: We conclude that D1-like receptors inhibit ROS production by altering PON2 distribution in membrane microdomains in the short-term, and by increasing PON2 expression in the long-term.
Keywords: NADPH oxidase; dopamine receptors; lipid rafts; paraoxonase 2; reactive oxygen species.
Figures
References
-
- Hussain T, Lokhandwala MF. Renal dopamine receptors and hypertension. Exp Biol Med. 2003;228:134–142. - PubMed
-
- Zeng C, Zhang M, Asico LD, Eisner GM, Jose PA. The dopaminergic system in hypertension. Clin Sci. 2007;112:583–597. - PubMed
-
- Beaulieu JM, Gainetdinov RR. The physiology, signaling, and pharmacology of dopamine receptors. Pharmacol Rev. 2011;63:182–217. - PubMed
-
- Asghar M, Tayebati SK, Lokhandwala MF, Hussain T. Potential dopamine-1 receptor stimulation in hypertension management. Curr Hypertens Rep. 2011;13:294–302. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous