

Animal models of Duchenne muscular dystrophy: from basic mechanisms to gene therapy
- PMID: 25740330
- PMCID: PMC4348559
- DOI: 10.1242/dmm.018424
Animal models of Duchenne muscular dystrophy: from basic mechanisms to gene therapy
Abstract
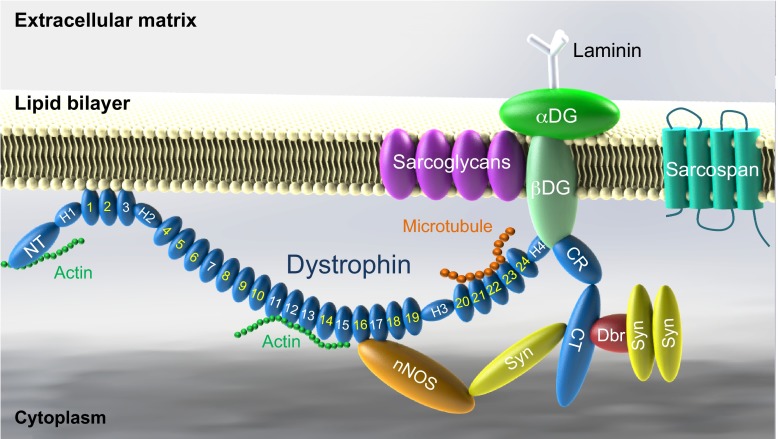
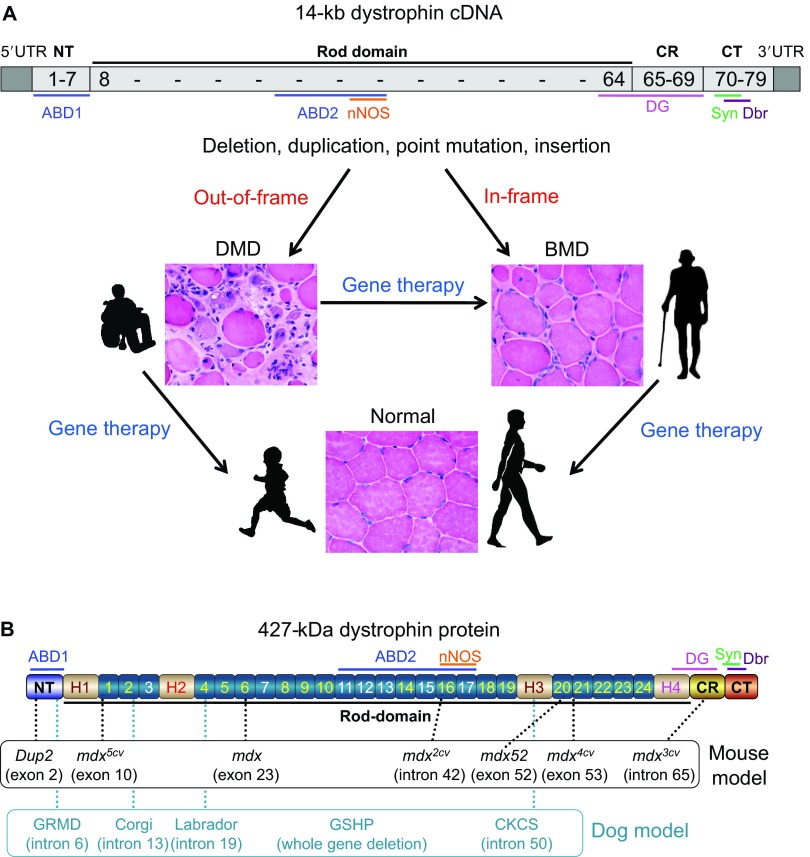
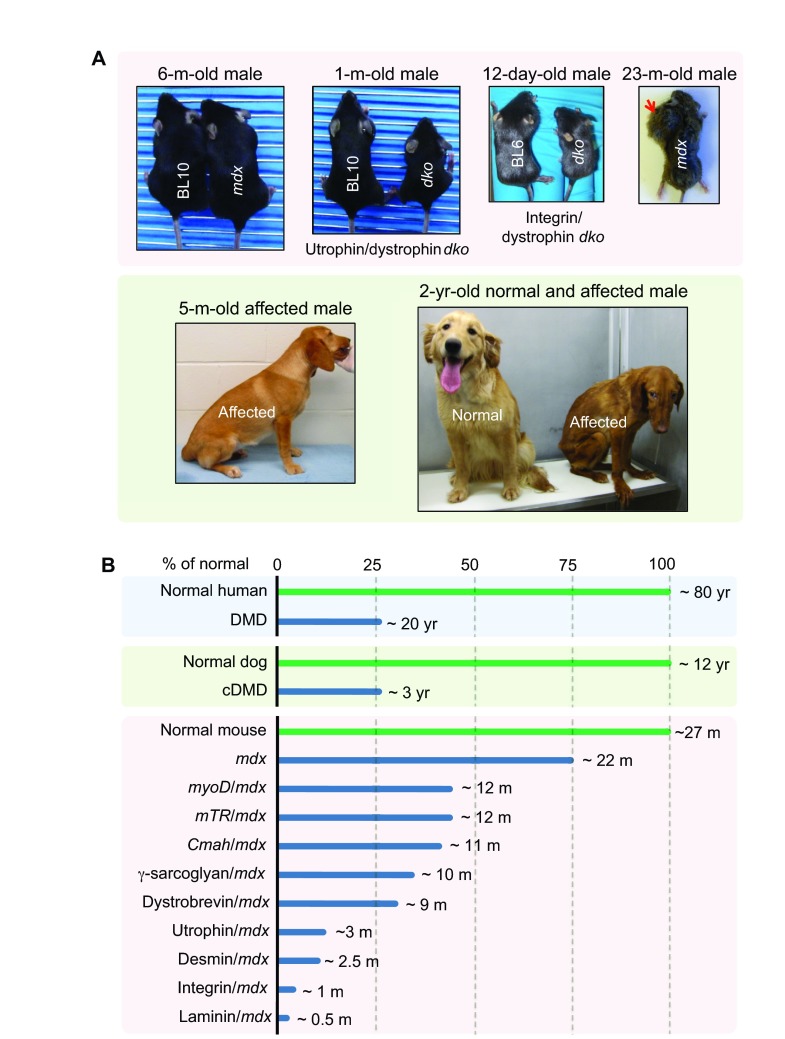
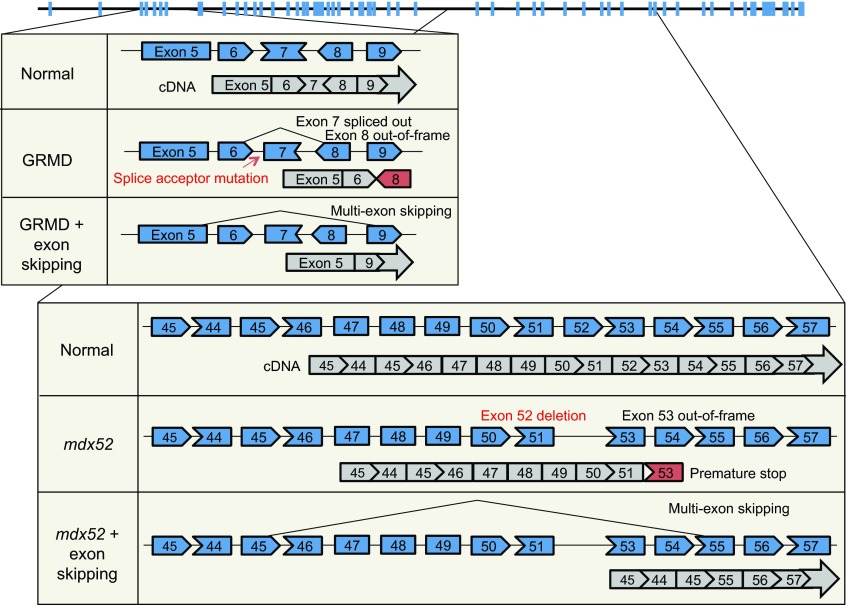
Duchenne muscular dystrophy (DMD) is a progressive muscle-wasting disorder. It is caused by loss-of-function mutations in the dystrophin gene. Currently, there is no cure. A highly promising therapeutic strategy is to replace or repair the defective dystrophin gene by gene therapy. Numerous animal models of DMD have been developed over the last 30 years, ranging from invertebrate to large mammalian models. mdx mice are the most commonly employed models in DMD research and have been used to lay the groundwork for DMD gene therapy. After ~30 years of development, the field has reached the stage at which the results in mdx mice can be validated and scaled-up in symptomatic large animals. The canine DMD (cDMD) model will be excellent for these studies. In this article, we review the animal models for DMD, the pros and cons of each model system, and the history and progress of preclinical DMD gene therapy research in the animal models. We also discuss the current and emerging challenges in this field and ways to address these challenges using animal models, in particular cDMD dogs.
Keywords: Animal model; Canine DMD; Duchenne muscular dystrophy; Dystrophin; Gene therapy.
© 2015. Published by The Company of Biologists Ltd.
Figures

References
-
- Aartsma-Rus A. (2012). Overview on DMD exon skipping. Methods Mol. Biol. 867, 97–116. - PubMed
-
- Aartsma-Rus A., Van Deutekom J. C., Fokkema I. F., Van Ommen G. J., Den Dunnen J. T. (2006). Entries in the Leiden Duchenne muscular dystrophy mutation database: an overview of mutation types and paradoxical cases that confirm the reading-frame rule. Muscle Nerve 34, 135–144. - PubMed
-
- Acsadi G., Dickson G., Love D. R., Jani A., Walsh F. S., Gurusinghe A., Wolff J. A., Davies K. E. (1991). Human dystrophin expression in mdx mice after intramuscular injection of DNA constructs. Nature 352, 815–818. - PubMed
-
- Ahn A. H., Kunkel L. M. (1993). The structural and functional diversity of dystrophin. Nat. Genet. 3, 283–291. - PubMed
-
- Aigner B., Rathkolb B., Klaften M., Sedlmeier R., Klempt M., Wagner S., Michel D., Mayer U., Klopstock T., de Angelis M. H., et al. (2009). Generation of N-ethyl-N-nitrosourea-induced mouse mutants with deviations in plasma enzyme activities as novel organ-specific disease models. Exp. Physiol. 94, 412–421. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
